

Un virus est un agent infectieux nécessitant un hôte, souvent une cellule, dont les constituants et le métabolisme déclenchent la réplication. Le nom virus a été emprunté au XVIe siècle par Ambroise Paré au latin vīrus, ī, n. (« venin, poison, proprement suc des plantes »). La science des virus est la virologie, et ses experts sont des virologues ou virologistes.
On considère de plus en plus les virus comme faisant partie des acaryotes. Ils changent de forme durant leur cycle, passant par deux stades :
- Une phase extracellulaire sous forme de particule virale. Ils sont alors des objets particulaires, infectieux, constitués au minimum d'un acide nucléique, souvent englobé dans une capside de protéines ;
- Une phase intracellulaire :
- soit comme séquence virale sous forme dormante,
- soit en train de détourner activement la machinerie cellulaire au profit de sa réplication en parasitant tout ou partie du métabolisme de son hôte.
Sous la forme intracellulaire (à l'intérieur de la cellule hôte), les virus sont des éléments génétiques qui peuvent s'intégrer à un chromosome du génome hôte (on parle alors de provirus ou de prophage) ou non (cas par exemple des usines à virions).
En 2022, 10 434 espèces de virus étaient décrites, réparties en 233 familles et 2 606 genres.
Pour les humains, seules 129 espèces sont jugées pathogènes en 2018.
Le débat sur la nature des virus (vivants ou pas) repose sur des notions complexes et reste aujourd'hui ouvert. Selon de nombreuses définitions du vivant (entité matérielle réalisant les fonctions de relation, nutrition, reproduction), les virus ne sont pas des êtres vivants, mais d'autres définitions permettent de les considérer comme vivants.
Découvertes
Identification des agents infectieux
Les maladies virales comme la rage, la fièvre jaune ou la variole affectent l'Homme depuis des millénaires. Des hiéroglyphes mettent en évidence la poliomyélite dans l'Égypte antique ; des écrits de l'Antiquité gréco-romaine et d'Extrême-Orient décrivent certaines maladies virales.
Varron (116–27 av. J.-C.) affirme "qu'il y a des créatures minuscules qui ne peuvent être vues par les yeux", qui flottent dans l'air et pénètrent dans le corps par la bouche et le nez et y provoquent des maladies graves.
À la fin du XIXe siècle, se représenter des agents infectieux qui ne fussent ni des bactéries, ni des champignons, ni des parasites, et qu'on ne pouvait déceler au microscope optique, était encore difficilement concevable. Le médecin testerin Jean Hameau avait fait un premier exposé sur les virus en 1837 devant la Société royale de médecine de Bordeaux, Réflexions sur les virus, puis devant l'Académie nationale de médecine en 1843. Son Mémoire sur les virus est présenté en séance de l'Académie de médecine le dimanche .
Les scientifiques isolaient alors les agents infectieux grâce aux filtres de porcelaine utilisés pour recueillir les bactéries. Entre 1887 et 1892, le botaniste russe Dimitri Ivanovski, en étudiant la mosaïque du tabac, montre que la sève des plantes malades contenait un agent infectieux non retenu par les filtres Chamberland (conçus par le biologiste du même nom). Ivanovski pensait à une toxine ou une très petite bactérie. C'est le chimiste hollandais Martinus Willem Beijerinck qui approfondit ces travaux et, en 1898, écarta à la fois l'hypothèse bactérienne, et l'hypothèse toxinique : diluant la sève de plantes infectées, il l'inocula à des plantes qui développèrent la maladie ; réitérant la manipulation, il put transmettre la maladie de multiples fois, montrant ainsi que la sève de la dernière plante infectée était aussi virulente que la première, effet qu'une toxine, après tant de dilutions n'aurait pu produire. Beijerinck appela l'agent Contagium vivum fluidum (« germe vivant soluble »).
À la même époque, le premier virus identifié est celui de la fièvre aphteuse, par Friedrich Löffler et Paul Frosch. Le premier virus pathogène de l'Homme identifié est celui de la fièvre jaune, entre 1900 et 1902. Louis Pasteur les nomma « infrabactéries », d'autres les qualifièrent de « virus filtrants » ou « virus ultrafiltrants ».
Étude et description
C’est pendant la Première Guerre mondiale que le Britannique Frederick Twort et le microbiologiste franco-canadien Félix d'Hérelle mettent en évidence le phénomène de « lyse transmissible » observable par la lyse des bactéries cultivées en milieu solide. Ce phénomène est dû à un virus de bactéries que Félix d'Hérelle qualifia de bactériophage. Les virus des plantes, des animaux, de l'Homme et des bactéries étaient ainsi découverts et leurs listes ne cessèrent de s'allonger au cours du XXe siècle.
Vers 1925, un virus était défini comme un « agent responsable d'une maladie infectieuse, parasite, de nature particulaire et de taille comprise entre 0,01 et 0,3 micromètre ».
L'apparition de la microscopie électronique dans les années 1930 permit l'observation des virus, mais on ne savait toujours pas à cette époque ce qu'ils étaient réellement. Le biochimiste américain Wendell Stanley cristallisa le virus de la mosaïque du tabac sous forme de cristal protéique en 1935. L'année suivante, des études complémentaires montrèrent que ce cristal contenait également de l'ARN. Les études ultérieures montrèrent que, selon les virus étudiés, ceux-ci étaient composés soit de protéines et d'ARN, soit de protéines et d'ADN. C'est en 1957 qu'André Lwoff proposa une définition claire et moderne des virus. En 1959, les microbiologistes Lwoff, Anderson et Jacob proposèrent le terme de virion pour définir la particule virale infectieuse
À partir des années 1960, les progrès des cultures cellulaires, de la microscopie électronique et de la biologie moléculaire permirent de progresser dans la compréhension des mécanismes de réplication des virus, dans la réalisation de diagnostics fiables et dans l'élaboration de vaccins.
Omniprésence des virus
On sait, depuis la fin du XXe siècle, que l'océan mondial est un immense réservoir de virus, de la surface aux évents hydrothermaux en passant par l'Arctique et les sédiments marins.
Dans l'eau de mer, la concentration en particules virales est de 106 à 108 particules par millilitre. En surface et près des rivages, les concentrations en virus habituellement rencontrées sont de l'ordre de 107 virus par millilitre (soit dix mille virus par millimètre cube (un millième de millilitre)) ; la concentration décroissant avec la profondeur et la distance au rivage. Des concentrations plus élevées (108 à 109 / cm3) se rencontrent dans les sédiments marins proches de la surface.
Ces virus jouent dans l'océan un rôle majeur dans le contrôle des efflorescences algales, ainsi que dans les cycles biogéochimiques, en particulier dans le cycle du carbone océanique (quotidiennement environ 20 % des organismes constituant la biomasse microbienne océanique totale est tuée par des virus ; ces derniers s'attaquent massivement au phytoplancton et au zooplancton, mais aussi aux bactéries et cyanophycées).
Grâce aux progrès de la cytométrie en flux et de l'analyse génétique (métagénomique notamment), en quelques décennies les chercheurs ont inventorié près de 200 000 types de populations virales en mer (en 2019, on en comptait 195 728 exactement, un chiffre douze fois plus élevé que celui de l'évaluation faite en 2016) ; 90 % des virus identifiés en mer entre 2016 et 2019 étaient jusqu'alors inconnus de la science. Remarque : on ne parle pas ici d'espèces mais de populations, au sein desquelles il y a plus de flux de gènes dans un groupe qu'entre groupes de virus (si les virus séquencés partagent au moins 95 % de leur ADN, alors ils sont classés dans une même population distincte des autres).
En 2007 on a estimé qu'il pourrait y avoir environ 1030 virus dans l'océan ; étirés et mis bout à bout, ils formeraient une ligne s'étendant au-delà des 60 galaxies les plus proches. Et chaque seconde il y aurait environ 1023 infections virales dans l'océan, jouant un rôle majeur dans l'évolution et l'entretien de la biodiversité marine. L'abondance virale semble liée à l'abondance et à la productivité en procaryotes, mais cette relation varie selon les environnements marins, notamment en fonction de la température.
Virome
Le virome est la composante virale d'un microbiome. Ainsi, le virome humain (en) est l'ensemble des communautés virales du microbiote de l'organisme humain. La recherche actuelle estime que dans le corps humain il y a 100 fois plus de virus (1015) que de cellules humaines (1013). Chaque individu en bonne santé porte en moyenne plus de 10 types de virus responsables d'infections virales systémiques chroniques et asymptomatiques.
Caractéristiques

On caractérise un virus par son incapacité à se reproduire par mitose, par scissiparité ou par méiose. Pour répliquer son acide nucléique, il dépend d'une cellule hôte qu'il doit infecter pour détourner et utiliser son métabolisme : un virus est nécessairement un parasite intracellulaire. Il est composé d'une ou plusieurs molécules d'acide nucléique (ADN ou ARN, simple ou double brin), éventuellement incluse dans une coque protéique appelée capside, voire d'une enveloppe lipidique (ex : l'Ebolavirus est un virus enveloppé). Parfois certaines capsides contiennent quelques enzymes (par exemple : transcriptase inverse du VIH) mais aucune pouvant produire de l'énergie.
Historiquement, les virus ont d'abord été considérés comme des particules organiques dites non filtrables, puis de petite taille (inférieure à celle d'une bactérie), en règle générale moins de 250 nanomètres, possédant un acide nucléique double ou simple toujours d'un seul type (ADN ou ARN). Les girus ont bousculé une première fois cette définition au moment de leur découverte. Ces derniers appartiennent pourtant bien au règne des virus et leurs virions possèdent à la fois des molécules d'ADN et d'ARN, remettant en cause cette vision historique. Il fallut repenser la définition des virus et la création de classes tels les « virus géants » comme mimivirus avec sa taille de 400 nm ou « girus » ou les NCLDV, voire les pandoravirus avec une taille allant jusqu'à 1 000 nm et leur « capside » qui n'en est pas vraiment une. La découverte des virophages et des virus satellites a aussi modifié la vision qu'on avait des virus, révoquant l'idée qu'une virose cellulaire était la forme irréductible du parasitisme.
Aujourd'hui les chercheurs s'accordent sur une remise en cause du paradigme capsidocentré, eu égard aux découvertes d'espèces virales montrant que certaines peuvent avoir plusieurs formes, y compris acapsidées, mais chaque fois infectieuses sans l'aide d'un virus assistant. Au-delà de ce paradigme, il semble que les origines des virus soient multiples. Ainsi certains virus auraient évolué à partir de putatifs ancêtres cellulaires s'étant simplifiés. Parallèlement, d'autres virus auraient évolué à partir de réplicons génétiques autonomes tels que les transposons, les plasmides et affiliés, finissant par acquérir d'abord une infectiosité propre puis une éventuelle capside.
Nature
S'ils sont inclus dans la biologie et l'étude des maladies, les virus sont l'objet de débats depuis leur première découverte et celles qui ont suivi.
Virus vivant ou pas
Le débat sur le caractère vivant ou inerte des virus reste encore aujourd'hui ouvert. Répondre à cette question exige de répondre au préalable à une autre : qu'est-ce que la vie ? D'après Ali Saïb, « la notion du vivant est une notion dynamique, évoluant en fonction de nos connaissances. En conséquence, la frontière entre la matière inerte et le vivant est tout aussi instable ». L'existence ou non d'un métabolisme, c'est-à-dire d'un ensemble cohérent de processus chimiques (l'homéostasie et non la reproduction), constitue un discriminant possible, en tout cas commode, mais qui semble réducteur.
Comme les cellules vivantes, les virus possèdent un acide nucléique (ADN ou ARN) et des protéines. Cependant, selon la définition du biochimiste Wendell Stanley, les virus ne sont pas des êtres vivants mais de « simples » associations de molécules biologiques, le fruit d'une auto-organisation de molécules organiques.
François Jacob insiste aussi sur cette caractéristique des virus : « Placés en suspension dans un milieu de culture, ils ne peuvent ni métaboliser, ni produire ou utiliser de l'énergie, ni croître, ni se multiplier, toutes fonctions communes aux êtres vivants. » Les virus n'ont pas leur propre machinerie enzymatique, ne peuvent se multiplier qu'en utilisant celle d'une cellule qu'ils infectent. Cependant ces caractéristiques sont en partie partagées avec les bactéries intracellulaire obligatoires.
En revanche, Gustavo Caetano-Anollés et Arshan Nasir (du laboratoire de bio-informatique évolutionnaire à l'université de l'Illinois, États-Unis) défendent une tout autre thèse. Ils avancent que, à côté des trois grandes « branches » du vivant (classiquement regroupées sous le nom de domaines) archées, bactéries (procaryotes) et eucaryotes, les virus en constitueraient une quatrième. Ils seraient la résultante de cellules ayant précédé le dernier ancêtre commun universel (Last Universal Common Ancestor, acronyme LUCA) des trois autres domaines. Pour avancer leur théorie les deux chercheurs se basent, non sur les séquences génétiques, mais sur les structures 3D des protéines qu'elles produisent.
Depuis 1990, ils ont analysé 11 millions de protéines produites par 3 460 espèces de virus et 1 620 espèces de cellules appartenant aux trois domaines ; ils affirment ainsi pouvoir retracer l'histoire évolutive de ces structures ; les protéines ayant des structures proches seraient issues d'un même ancêtre hypothétique.
Si cette hypothèse est encore minoritaire, Patrick Forterre, biologiste spécialiste de l'évolution, considère qu'elle a le mérite de « favoriser le retour des virus dans la mire des évolutionnistes, alors qu'ils en étaient les grands absents ».
En élargissant la définition des virus aux cellules virolées comme étant transformées en cellules virales, et les particules virales (en particulier le virion avant changement de paradigme) comme des éléments reproducteurs plutôt que le virus lui-même (à l'image de spores), alors il est acceptable selon ce point de vue d'envisager les virus comme vivants.
Des entités intermédiaires
- Mimivirus
Au cours des dernières années, des entités intermédiaires ont été découvertes : le mimivirus, infectant une amibe, possède dans son génome 1 200 gènes (davantage que certaines bactéries). Certains de ces gènes participeraient à la synthèse protéique et à des mécanismes de réparation de l'ADN. Il existe chez le mimivirus une trentaine de gènes présents habituellement chez les organismes cellulaires mais absents chez les virus.
- Virus ATV
Le virus ATV d'archées présente lui aussi des caractéristiques étonnantes : ce virus en forme de citron présente la particularité de se modifier en dehors du contexte cellulaire par un mécanisme actif. Il est capable de s'allonger à chaque extrémité à une température de 80 °C, température à laquelle vit son hôte Acidianus à proximité des sources hydrothermales. Néanmoins, organes et échanges cycliques, donc métabolisme, restent absents.
Virus et évolution
Les virus jouent aussi un rôle dans l'évolution. Patrick Forterre avance même l'hypothèse que les virus seraient les premiers organismes à ADN. À l'origine de la vie, l'ARN dominait (hypothèse du monde à ARN) et assurait à la fois les fonctions de stockage et transmission de l'information génétique et de catalyse des réactions chimiques. Seules existaient des cellules dont le génome était codé par de l'ARN et dont le métabolisme était assuré par des ARN-enzymes qui ont progressivement été remplacés par des protéines-enzymes. Ces protéines, déjà complexes, auraient « inventé » l'ADN. L'ADN a été sélectionné en raison de sa plus grande stabilité. D'après Patrick Forterre, l'ADN confèrerait au virus le pouvoir de résister à des enzymes dégradant les génomes à ARN, arme de défense probable des protocellules. On retrouve le même principe chez des virus actuels, qui altèrent leur ADN pour résister à des enzymes produites par des bactéries infectées.
Virus et microbes
Virus et micro-organismes (ou microbes) ne sont donc pas des notions de même nature. Elles s'opposent en ce que les microbes sont des organismes vivants, ce qui est contesté pour les virus. Mais leur portée est différente, les micro-organismes (bactéries, archées, levures, protistes, etc.) n'étant regroupés que pour leur taille microscopique, sans que ce regroupement ait un sens en termes de classification des espèces, alors que les virus ont bien des caractéristiques phylogéniques communes, même si le concept d'espèce reste flou pour les acaryotes.
Structure
Tout agent infectieux ressortissant au règne des virus est composé au minimum d'un acide nucléique. Les formes incapables d'effectuer le cycle viral sans assistance sont qualifiées particules sub-virales (exemple : virusoïde, ADN satellite, etc.). Les formes extracellulaires capables d'effectuer le cycle viral sans assistance sont appelées particules virales, allant d'une forme simplifiée à l'extrême et ne comportant que l'acide nucléique — lequel, lorsqu'il encode au minimum une protéine, est qualifié de virus et lorsqu'il n'encode aucune protéine est appelé viroïde — ou une forme transportant un à plusieurs acides nucléiques dans un conteneur protéique appelée virion.
On le dit encapsidé car l'acide nucléique, généralement stabilisé par des nucléoprotéines basiques, est enfermé dans une coque protéique protectrice appelée capside. La forme de la capside est à la base des différentes morphologies des virus. Le virion a une forme microscopique variable : si la représentation « usuelle » lui donne l'image du VIH, les différentes espèces ont des formes allant de la sphère à des conformations d'apparence insectoïde.
La taille des virus se situe entre 10 et 400 nanomètres. Les génomes des virus ne comportent que de quelques gènes à 1 200 gènes. L'un des plus petits virus connus est le virus delta, qui parasite lui-même celui de l'hépatite B. Il ne comporte qu'un seul gène. L'un des plus gros virus connus est le mimivirus, avec un diamètre qui atteint 400 nanomètres et un génome qui comporte 1 200 gènes.
Acide nucléique
Le filament d'acide nucléique peut être de l'ADN ou de l'ARN. Il représente le génome viral. Il peut être circulaire ou linéaire, bicaténaire (double brin) ou monocaténaire (simple brin). Le génome sous forme d'ADN est généralement bicaténaire. Le génome sous forme d'ARN est généralement monocaténaire et peut être à polarité positive (dans le même sens qu'un ARN messager) ou à polarité négative (complémentaire d'un ARN messager). Le peloton central d'acide nucléique est dénommé nucléoïde.
Capside
La capside est une coque qui entoure et protège l'acide nucléique viral. La capside est constituée par l'assemblage de sous-unités protéiques appelées capsomères. L'ensemble formé par la capside et le génome est nommé nucléocapside. La structure de la capside peut présenter plusieurs formes. On distingue en général deux groupes principaux de virus : les virus à symétrie cubique (ou à capside icosaédrique) et les virus à symétrie hélicoïdale.
Enveloppe
De nombreux virus sont entourés d'une enveloppe (ou péplos) qui prend naissance au cours de la traversée des membranes cellulaires. Sa constitution est complexe et présente un mélange d'éléments cellulaires et d'éléments d'origine virale. On y trouve des protéines, des glucides et des lipides. Les virus possédant une enveloppe sont les virus enveloppés. Les virus ne possédant pas d'enveloppe sont les virus nus. L'enveloppe permet un camouflage immunologique qui est un avantage évolutif, en revanche les virus enveloppés sont plus sensibles que les virus nus à la désinfection par des détergents doux.
| Virus icosaédriques | |
| La capside icosaédrique entraîne une apparence sphérique du virus. Les protomères sont organisés en capsomères, disposés de manière régulière et géométrique. Un capsomère est composé de cinq ou six protomères, appelés pentons aux sommets et hexons au niveau des faces et des arêtes. Parmi les virus icosaédriques, les parvovirus ont une capside formée de 12 capsomères, les poliovirus 32 capsomères, les papillomavirus 72 capsomères, tandis que la capside des adénovirus est constituée de 252 capsomères. |
|
| Virus hélicoïdaux | |
| Ces virus sont de longs cylindres (300 à 400 nm), creux, composés d'un type de protomère enroulé en spirale hélicoïdale formant des anneaux appelés capsomères. Ils peuvent être rigides ou flexibles. Le matériel génétique est logé à l'intérieur du tube. Le virus de la mosaïque du tabac est un exemple de virus hélicoïdal très étudié. | |
| Virus enveloppés | |

Schéma d'un virus enveloppé : le VIH.
|
En plus de la capside, certains virus sont capables de s'entourer d'une structure membranaire empruntée à la cellule hôte. Cette enveloppe membranaire est composée d'une bicouche lipidique qui peut posséder des protéines codées par le génome viral ou le génome de l'hôte. Cette enveloppe donne quelques avantages aux virions par rapport à ceux composés d'une capside seule, comme la protection vis-à-vis d'enzymes ou de composés chimiques. Les virus enveloppés sont par contre plus fragiles dans l'environnement extérieur, sensibles aux détergents et à la dessiccation. Les glycoprotéines, formant des spicules, fonctionnent comme des récepteurs permettant de se fixer sur des cellules hôtes spécifiques. Les virus de la grippe (famille des Orthomyxoviridae), le VIH (famille des Retroviridae), sont des exemples de virus enveloppés. |
| Virus complexes | |

Schéma d'un bactériophage.
|
Ces virus possèdent une capside symétrique qui n'est ni hélicoïdale, ni vraiment icosaédrique. Les bactériophages comme le phage T4 d’Escherichia coli sont des virus complexes possédant une tête icosaédrique liée à une queue hélicoïdale à laquelle sont attachés des poils et des fibres caudales. Le poxvirus (variole, vaccine) est aussi un exemple de virus complexe. C'est un des plus grands virus animaux (250 à 350 nm de long sur 200 à 250 nm de large). Certains virus se présentent sous une forme bacillaire. C'est le cas du virus de la rage (famille des Rhabdoviridae) et du virus Ébola. |

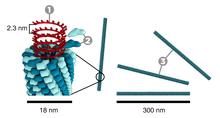
Réplication
Différentes voies
On distingue deux voies principales de réplication du génome viral :
- le cycle lytique, considéré comme la réplication virale dans sa phase active ;
- le cycle lysogénique, considéré comme la "phase dormante" du virolage.
Multiplication virale
Les virus ne peuvent se répliquer qu'au sein de cellules vivantes. C'est l'interaction du génome viral et de la cellule hôte qui aboutit à la production de nouvelles particules virales. L'infection d'une cellule par un virus, puis la multiplication du virus, peuvent se résumer en différentes étapes. Toutefois, après pénétration du virus dans la cellule, ces étapes peuvent différer selon la nature du virus en question et notamment selon qu'il s'agit d'un virus à ADN ou d'un virus à ARN, ou encore d'un girus.
- Adsorption du virus au contact de la membrane de la cellule infectée, grâce à des récepteurs spécifiques
- Pénétration dans la cellule
- Décapsidation (libération de l'acide nucléique)
- Réplication du génome viral
- Biosynthèse des protéines virales
- Assemblage et encapsidation des particules virales produites
- Libération des virions hors de la cellule-hôte
- Structures réplicatives remarquables
Certains virus induisent des structures où se concentrent l'activité réplicative :
Culture des virus
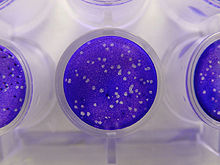
Afin de mieux connaître la biologie, la multiplication et le cycle des virus, et éventuellement de préparer des vaccins, il est nécessaire de cultiver les virus. Ceux-ci peuvent se multiplier uniquement au sein de cellules vivantes. Les virus infectant les cellules eucaryotes sont cultivés sur des cultures de cellules obtenues à partir de tissus animaux ou végétaux. Les cellules sont cultivées dans un récipient en verre ou en plastique, puis sont infectées par le virus étudié. Les virus animaux peuvent aussi être cultivés sur œufs embryonnés et parfois chez l'animal, lorsque la culture in vitro est impossible. Les virus bactériens peuvent également être cultivés par inoculation d'une culture bactérienne sensible. Les virus de végétaux peuvent aussi être cultivés sur des monocouches de tissus végétaux, des suspensions cellulaires ou sur des plantes entières.
Les virus peuvent ensuite être quantifiés de différentes manières. Ils peuvent être comptés directement grâce à la microscopie électronique. Dans le cas des virus bactériens, la technique des plaques virales (ou plages) est très utilisée pour évaluer le nombre de virus dans une suspension. Une dilution de suspension virale est ajoutée à une suspension bactérienne, puis l'ensemble est réparti dans des boîtes de Petri. Après culture, des zones claires (plages) à la surface de la gélose sont la conséquence de la destruction d'une bactérie et des bactéries adjacentes par un virion.
Les virus peuvent être purifiés grâce à diverses méthodes de biochimie (centrifugation différentielle, précipitation, dénaturation, digestion enzymatique).
Origine
Tout être vivant peut être infecté par un virus. Il existe des virus de bactéries (les bactériophages), des virus d'archées, des virus d'algues (Phycodnaviridae), des virus de plantes, des virus fongiques, des virus d'animaux, parmi lesquels on trouve de nombreux agents pathogènes, et même des virus de virus.
Il existe plusieurs hypothèses concernant l'origine et l'évolution des virus. Il est probable que tous les virus ne dérivent pas d'un ancêtre commun et les différents virus peuvent avoir des origines différentes.
- Les virus et les cellules ont pu apparaître dans la soupe primordiale en même temps et évoluer parallèlement. Dans ce scénario, au début de l'apparition de la vie, les plus anciens systèmes génétiques d'auto-réplication (probablement de l'ARN) sont devenus plus complexes et se sont enveloppés dans un sac lipidique pour aboutir au progénote à l'origine des cellules. Une autre forme réplicative aurait pu garder sa simplicité pour former des particules virales.
- Les virus pourraient avoir pour origine des morceaux d'acides nucléiques qui se sont « échappés » du génome cellulaire pour devenir indépendants. Ce phénomène pourrait avoir eu lieu lors d'erreurs au cours de la réplication du matériel génétique. Les virus pourraient aussi avoir pour origine des plasmides (molécules d'ADN circulaires) ou des transposons (séquences d'ADN capables de se déplacer et de se multiplier dans un génome), voire des viroïdes.
- Les virus pourraient dériver de cellules ayant subi une simplification. D'après cette hypothèse, les ancêtres des virus auraient été des êtres vivants libres ou des micro-organismes devenus des prédateurs ou des parasites dépendants de leur hôte. Les relations de parasitisme entraînent la perte de nombreux gènes (notamment les gènes pour le métabolisme apportés par l'hôte). Cet organisme aurait coévolué avec la cellule hôte et n'aurait conservé que sa capacité à répliquer son acide nucléique et le mécanisme de transfert de cellule à cellule. Cette hypothèse s'appuie notamment sur l'existence des rickettsies, petites bactéries ayant régressé à un tel point qu'elles ne peuvent survivre que dans une cellule hôte, et rappelant les virus.
Des études en 2013 de divers girus tendent à favoriser l'hypothèse d'une simplification. Cela impliquerait que les virus pourraient être un embranchement phylogénétique au même titre que les autres règnes (eucaryotes, bactéries, archées) du vivant.
Rôle dans l'évolution
Il est possible que les virus soient très anciens, peut-être plus anciens que les bactéries les plus âgées.
Au début des années 2000, dans des amibes du genre Acanthamoeba, des chercheurs ont découvert un virus géant (Megaviridae) : le Mimivirus. Aussi grand et complexe que certaines bactéries, il a modifié la perception des virologistes quant aux limites supérieures de taille (sa longueur totale dépasse 0,7 micromètre) et du nombre de gènes du monde viral (il possède plus de 1 000 gènes).
Dix ans plus tard, des chercheurs français publiaient (2013) la description de deux virus encore plus grands, et dont le génome est environ deux fois plus gros (en nombre de gènes) que les précédents virus géants découverts. Ces deux nouveaux virus géants ont été classés dans une catégorie créée pour eux (Pandoravirus) car ils ne sont pas apparentés aux virus connus et présentent même des caractéristiques inattendues :
- leur diamètre approche le micromètre et dépasse le record de Megavirus chilensis ;
- leur génome a une taille très supérieure à ce qui était connu : environ 2 500 gènes ; à titre de rappel, le génome de virus tels que ceux de la grippe ou de l'immunodéficience humaine ne contiennent qu'une dizaine de gènes ;
- leur génome ne code qu'une infime part (6 %) des protéines habituellement produites par les autres virus connus ;
- ils ne disposent pas des gènes nécessaires à la synthèse de la protéine de capside (la « brique de base » des capsides de virus normaux). L'analyse du protéome de Pandoravirus salinus a confirmé que les protéines qui le constituent sont bien celles que l'on peut prédire à partir de la séquence génomique virale.
Le premier (Pandoravirus salinus) a été trouvé dans des sédiments marins prélevés au large du Chili et le second (Pandoravirus dulcis) dans une mare d'eau douce près de Melbourne (en Australie).
Bien que présentant les caractères essentiels d'un virus (pas de ribosome, pas de division ni de production d'énergie), ils semblent d'un type tout à fait nouveau. Leur génome dépasse en taille celui de certains petits eucaryotes (cellules à noyau) parasites.
Les Pandoravirus utilisent donc directement le code génétique de leur hôte. Ces organismes ne sont pourtant ni des eucaryotes, ni des eubactéries ni des archébactéries. Cette découverte remet en question le dogme établi par la virologie dans les années 1950 voulant qu'il n'y ait pas de continuité entre virus et bactéries. La vie cellulaire aurait donc pu émerger à partir de formes de vie pré-cellulaires plus variées que ce qu'on pensait.
D'autre part, les virus jouent un rôle important de vecteur naturel dans les transferts de gène dits horizontaux (par opposition aux transferts dits verticaux de parent à descendant) entre différents individus et même différentes espèces, permettant un accroissement de diversité génétique, et la dissémination d'innovations génétiques au-delà de la descendance d'individu porteur d'une mutation génétique donnée. En particulier la transduction et l'endogénisation sont typiquement des évolutions génétiques qui ne peuvent s'effectuer qu'à l'aide des virus.
En abiotique (les prémisses du vivant), une des hypothèses stipule que les virus auraient joué des rôles clefs très tôt dans l'histoire évolutive du vivant, probablement avant la divergence entre bactéries, archées et eucaryotes, à l'époque du dernier ancêtre commun universel. Ils restent l'un des plus grands réservoirs de diversité génétique inexplorés sur la planète.
Les virus jouent également un rôle important dans les corps des humains. Selon le chercheur Clément Gilbert, « le corps d'un homme adulte sain abrite plus de trois mille milliards de virus, pour la plupart des bactériophages infectant les bactéries présentes dans le tractus intestinal et sur les muqueuses. L'impact de ces virus n'est pas encore complètement compris, mais on peut déjà parier qu'ils jouent un rôle important dans la régulation de la composition des communautés bactériennes vivant en symbiose avec l’homme ». Il souligne également que « plus de 8 % du génome humain dérivent de rétrovirus », ce qui permet de dire que « nous sommes d’une certaine manière apparentés aux virus ».
Virus et maladies
Les virus possèdent différents mécanismes leur octroyant diverses possibilités stratégiques d'infection, dont l'incidence provoque éventuellement des maladies. Le virion pénètre une cellule hôte plus ou moins spécifique où il se désagrège, libérant son contenu qui en s'activant prend le pas sur les fonctions cellulaires normales. À ce niveau, les effets cytopathogènes des virus peuvent entraîner divers effets néfastes. Les capacités de synthèse protéique de la cellule infectée peuvent être détournées ou inhibées, tandis que la chromatine est fragmentée par des enzymes virales. Des particules virales s'accumulent dans le cytoplasme avant de s'assembler en virions. La surcharge virale endo-cellulaire provoque enfin la mort de la cellule hôte par lyse, libérant les virions qui vont ensuite disséminer.
Lorsque le virus pénètre dans une cellule non permissive, il ne peut pas se multiplier. Son génome peut cependant subsister sous la forme d'un épisome libre ou intégré au génome cellulaire. Il y a transformation cellulaire virale lorsque le génome du virus entre en interaction avec l'ADN du génome cellulaire. On appelle ces virus des virus oncogènes. Parmi ceux-ci, les rétrovirus, en s'intégrant dans le génome cellulaire, peuvent devenir tumorigènes et éventuellement entraîner des cancers.
La capacité d'un virus d'entraîner une maladie est décrite en termes de pouvoir pathogène tandis que son intensité est exprimée en termes de virulence. La classification des principaux groupes de virus, et leurs correspondances en pathologie, se trouvent dans l'encyclopédie médicale Vulgaris. Cette classification est notamment basée sur le type de molécules d'acide nucléique (ARN ou ADN) dont est constitué le virion.
Maladies humaines


En 2018, on recense 129 espèces de virus impliqués dans des maladies humaines.
Le rhume, la grippe, la varicelle, la rougeole, la mononucléose infectieuse sont des exemples de maladies humaines relativement courantes d'origine virale. On connaît d'autres exemples plus nocifs comme le SIDA, certains coronavirus (SRAS, maladie à coronavirus 2019), la grippe aviaire, la variole, ou la maladie à virus Ebola, fièvre hémorragique causée par le virus Ebola.
Quelques exemples de virus pathogènes pour Homo sapiens :
- VIH, virus du SIDA
- Virus coxsackie A
- Virus Ebola
- Virus de la poliomyélite
- Virus de la variole
- Virus de la grippe
- Virus de la fièvre jaune
- Virus du Nil occidental
- Cytomégalovirus
- Rotavirus
- Virus de l'Hépatite C
- Virus simien 40 ou SV40
- Coronavirus (SRAS, MERS et COVID-19)
Dangerosité d'un virus : selon le professeur Arnaud Fontanet, épidémiologiste, qui dirige l'unité d'épidémiologie des maladies émergentes à l'institut Pasteur de Paris, les caractéristiques d'un virus dangereux :
- transmission respiratoire ;
- taux de reproduction de base supérieur à 2 ;
- taux de mortalité supérieur à 1/1000 ;
- temps de génération inférieur à trois jours ;
- contagion avant l'apparition des symptômes.
Prévention et traitements

Étant donné que les virus utilisent la machinerie cellulaire de l'hôte pour se reproduire à l'intérieur même de la cellule, il est difficile de les éliminer sans tuer la cellule hôte. Des médicaments antiviraux permettent cependant de perturber la réplication du virus.
Une autre approche est la vaccination qui permet de résister à l'infection.
Divers médicaments permettent de traiter les symptômes liés à l'infection, mais pas les antibiotiques, qui sont sans effet sur les virus. Les antibiotiques interfèrent en effet avec des constituants ou le métabolisme des bactéries et permettent donc de traiter seulement les maladies d'origine bactérienne et non les maladies d'origine virale.
Diverses méthodes de désinfection in vitro permettent d'inactiver les virus (hypochlorite de sodium à 1 %, éthanol à 70 %, glutaraldéhyde à 2 %, formaldéhyde, eau oxygénée à 2 %, acide peracétique).
Biotechnologie
Les virus présentant en général un matériel génétique simpliste, ce sont d'excellents outils dans l'étude de la biologie moléculaire et la biologie cellulaire. Ils permettent la manipulation de fonctions cellulaires, ce qui permet d'en approfondir notre compréhension et d'élucider certains mécanismes moléculaires de la génétique comme la réplication de l'ADN, la transcription, les modifications post-transcriptionnelles de l'ARN, la traduction, le transport des protéines et l'immunologie.
Les virus peuvent être utilisés (virothérapie) comme vecteur de gène au sein de cellules cibles. Outil utilisé par exemple pour faire acquérir à une cellule la capacité de produire une protéine d'intérêt ou pour étudier l'effet de l'introduction du nouveau gène dans le génome.
Certains virus sont utilisés en thérapie génique pour soigner diverses maladies génétiques, par exemple pour remplacer un gène défectueux provoquant des troubles fonctionnels ou mécaniques.
Les virus sont également utilisés dans la lutte contre le cancer. Certains virus peuvent être en quelque sorte programmés pour détruire spécifiquement des cellules cancéreuses.
Classification
Les virus sont classés selon leur stratégie de réplication, c'est-à-dire selon la nature de l'acide nucléique de leur génome (ADN ou ARN), la structure de l'acide nucléique (monocaténaire ou bicaténaire) et la forme de l'acide nucléique (linéaire, circulaire, segmenté ou non) : c'est la classification de Baltimore. Les données morphologiques peuvent également être prises en compte (présence ou absence d'enveloppe, symétrie de la capside). Souvent, le sérogroupage est encore utilisé pour raffiner la définition des différences entre virus très proches.
Un pas vers une classification phylogénétique est franchi en octobre 2018 avec la reconnaissance par l'ICTV (Comité international de taxonomie des virus) du regroupement des virus à ARN simple brin à polarité négative en un embranchement, deux sous-embranchements et six classes.
Virus de procaryotes
Il existe deux catégories de virus de procaryotes selon le type d'hôte qu'ils parasitent. La première catégorie regroupe ceux qui infectent les bactéries et sont appelés bactériophages. La deuxième catégorie regroupe ceux qui infectent les archées. Il existe quatre grands groupes morphologiques de virus de procaryotes.
- Les virus à symétrie binaire. Ce groupe représente près de 96 % des virus de procaryotes et correspond aux familles des Myoviridae, des Siphoviridae et des Podoviridae.
- Les virus à symétrie cubique avec une capside icosaédrique mais pas de queue comme les Microviridae.
- Les virus à symétrie hélicoïdale qui ont une forme de filaments comme les Inoviridae comme le phage M13.
- Les virus pléomorphes, sans capside véritable mais possédant une enveloppe. Ce groupe rassemble six familles de virus dont cinq regroupent des virus infectant seulement les archées. Certains virus d'archées sont pléomorphes, alors que d'autres ont des formes de bouteille, de citron, de fuseau.
Les bactériophages possèdent un rôle dans les écosystèmes. Par exemple, dans les écosystèmes aquatiques, ils participent au contrôle de l'abondance et de la diversité bactérienne.
Virus d'eucaryotes
Virus d'animaux
Virus des vertébrés
En principe spécifiques d'une espèce ou d'un groupe de phylums génétiquement proches, les virus ont tendance à infecter un type cellulaire ou tissulaire principal ou exclusif. Cependant, il existe de nombreux virus, comme la rage, qui sont moins spécifiques à un hôte par comparaison avec d'autres virus comme la maladie de Carré, le virus de l'immunodéficience féline ou la variole. Les virions se propagent principalement par contact direct entre individus, mais peuvent aussi diffuser dans l'air sous forme d'aérosols (éternuements), être charriés par des excrétions diverses (vomis, urines, selles, larmes…), ou encore transportés par d'éventuels arthropodes parasites (moustiques, tiques, puces…).
Virus des arthropodes
Les arbovirus sont des virus ayant pour vecteurs des arthropodes hématophages.
Les baculovirus sont des virus d'insectes très étudiés. Ils infectent principalement les lépidoptères. La larve de l'insecte s'infecte en ingérant de la nourriture. À partir du tube digestif, l'infection peut se transmettre aux autres tissus. L'utilisation de virus pathogènes d'invertébrés dans la lutte contre les insectes ravageurs des cultures et des forêts pourrait être l'un des moyens pour limiter ou remplacer les insecticides chimiques.
Les baculovirus sont aussi utilisés en biologie moléculaire pour exprimer un gène étranger (protéine recombinante) dans des cultures de cellules d'insectes.
Par ailleurs, certains virus de végétaux sont transmis par des invertébrés mais ne se multiplient pas chez ces vecteurs.
Virus des plantes
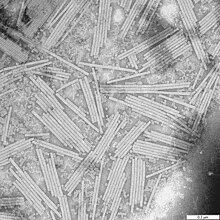
La structure des virus des plantes ou phytovirus, est similaire à celle des virus bactériens et animaux. Beaucoup de virus végétaux se présentent sous la forme de minces et longues hélices. La majorité a un génome composé d'ARN. Les virus de végétaux peuvent être disséminés par le vent ou par des vecteurs comme les insectes et les nématodes, parfois par les graines et le pollen. Les virus peuvent aussi contaminer la plante par l'intermédiaire d'une blessure ou d'une greffe.
Différents types de symptômes peuvent apparaître sur la plante infectée. Les virus peuvent provoquer des taches ou des flétrissements sur les feuilles et les fleurs. Des tumeurs peuvent survenir sur les tiges ou les feuilles.
Le virus de la mosaïque du tabac (TMV ou tobamovirus) est un exemple très étudié de virus de végétaux.
Virus des mycètes
Les virus des champignons, ou mycovirus, sont particuliers car ils se propagent lors de la fusion cellulaire. Il n'y a pas de virions extracellulaires. Chez les levures comme Saccharomyces, les virus sont transmis au moment du brassage cytoplasmique lors de la fusion cellulaire. Les champignons filamenteux comme Penicillium ou le champignon de Paris Agaricus bisporus peuvent également être infectés par des virus, ce qui peut entraîner des problèmes lors de production. Il a été imaginé d'utiliser ces virus dans le cadre d'une lutte biologique contre des champignons pathogènes.
Virus de virus
Découvert en 2008, Sputnik est un cas à part capable d'infecter un autre virus (Mamavirus) appartenant à la classe des virus géants (génome de plus de 300 000 pb et taille supérieure à 0,2 μm).
On connaît aussi d'autres virophages comme Mavirus associé au CroV (en) (un virus géant infectant l'hôte eucaryote Cafeteria roenbergensis, organisme unicellulaire).
Notes
Références
Voir aussi
Bibliographie
- Pierre Ardoin : Virus et diagnostic virologique. Paris, Maloine Éditeur, 1983, 997 p. ;
- Patrick Berche, « Vers des armes biologiques de nouvelle génération », Politique étrangère, vol. 70e année, no 1, , p. 137-146 (lire en ligne, consulté le ) ;
- Thierry Borrel : Les Virus. Diversité et organisation du monde viral. Interactions avec le vivant. Nathan Université, Paris, 1996 ;
- Jeanne Brugère-Picoux, « Passage de la barrière d’espèce », Les Cahiers du Musée des Confluences, t. 6, , p. 55-64 (lire en ligne, consulté le ) ;
- (en) Dimmock NJ, Easton AJ, Leppard K : Introduction to Modern Virology, 6th Ed. Blackwell Publishing, Oxford, UK, 2007 ;
- Gessain A., Manuguerra J.C. : Les virus émergents. Collection « Que sais-je ? », Presses Universitaires de France, 2006 ;
- (en) Madigan, M. T., Martinko, J. M. : Brock Biology of Microorganisms, 11th Ed. Pearson Prentice Hall, Upper Saddle River, NJ, 2005 ;
- Perry J., Staley J., Lory S. : Microbiologie. Éditions Dunod, 2004 ;
- Prescott, L.M., Harley, J.P. Klein, D.A. : Microbiologie 2e édition. DeBoeck eds, 2003 ;
- Saluzzo JeaN-François, La guerre contre les virus, Plon, 2002 ;
- Tania Louis (postface Étienne Klein), La folle histoire des virus, Paris, Humenscience, coll. « Comment a-t-on su ? », , 353 p. (ISBN 978-2-37931-194-9).
- (en) Roya Zandia, Bogdan Dragne, Alex Travesset et Rudolf Podgornik, « On virus growth and form », Physics Reports, vol. 847, , p. 1-102 (DOI 10.1016/j.physrep.2019.12.005) ;
- Carl Zimmer (trad. de l'anglais par Karim Madjer et Alan Vonlanthen), Planète de virus [« A planet of viruses »], Paris, Éditions Belin, coll. « Science à plumes », , 125 p. (ISBN 978-2-7011-9767-8) ;
Articles connexes
Liens externes
- Ressources relatives à la santé :
- (en) Medical Subject Headings
- (en) NCI Thesaurus
- (no + nn + nb) Store medisinske leksikon
- (cs + sk) WikiSkripta
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :