

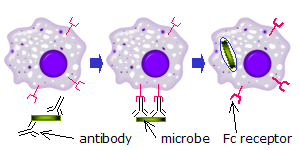
Un récepteur Fc est une protéine transmembranaire présente à la surface de certaines cellules —dont les lymphocytes B, les cellules dendritiques , les lymphocytes NK, les macrophages, les neutrophiles, les éosinophiles, les basophiles, les thrombocytes et les mastocytes — qui contribuent aux fonctions protectrices du système immunitaire. Les récepteurs Fc se lient aux anticorps qui ont opsonisé (c'est-à-dire « enrobé ») les pathogènes tentant d'envahir des cellules. Leur activité stimule la destruction des microbes par des cellules phagocytes ou cytotoxiques, ou celle des cellules infectées par phagocytose ou cytotoxicité à médiation cellulaire dépendante des anticorps.
Leur nom est dérivé de leur liaison spécifique à une zone des anticorps connue sous le nom de région Fc (Fragment cristallisable). Les récepteurs Fc font le pont entre l'immunité humorale du système immunitaire adaptatif représenté par les anticorps et les cellules du système immunitaire inné.
L'action stimulatrice ou inhibitrice sur la réponse immunitaire des récepteurs Fc peut être perturbée et déclencher plusieurs maladies auto-immunes.
Certains virus tels que les flavivirus utilisent les récepteurs Fc pour infecter les cellules, par un mécanisme connu sous le nom de facilitation dépendante des anticorps.
Fonctions
Les récepteurs Fc sont présents sur nombre de cellules du système immunitaire, dont les phagocytes tels que macrophages et monocytes ; les granulocytes tels que neutrophiles, éosinophiles et lymphocytes du système immunitaire inné (lymphocytes NK) ou du système immunitaire adaptatif (par exemple, lymphocytes B). Ils permettent à ces cellules de se lier aux anticorps qui sont attachés à la surface des microbes ou aux microbes de cellules infectées, en aidant ces cellules à identifier et à éliminer les microbes pathogènes. Les récepteurs Fc lient les anticorps à leur région Fc, une interaction qui active la cellule possédant le récepteur Fc. L'activation des phagocytes est la fonction la plus communément attribuée aux récepteurs Fc. Par exemple, les macrophages commencent à ingérer et à tuer les agents pathogènes enveloppés d'IgG en les phagocytant quand ils se sont liés à leurs récepteurs Fcy. Un autre processus impliquant les récepteurs Fc est appelé cytotoxicité à médiation cellulaire dépendante des anticorps (ADCC). Au cours de l'ADCC, les récepteurs FcγRIII présents sur la surface des lymphocytes NK stimulent la libération de molécules cytotoxiques à partir de leurs granules par les cellules NK, pour tuer les cellules cibles couvertes d'anticorps.
FcRI a une fonction différente. FceRI est le récepteur Fc des granulocytes, qui est impliqué dans les réactions allergiques et de défense contre les parasites infectieux. Lorsqu'un certain allergique de l'antigène ou du parasite est présent, la réticulation d'au moins deux des IgE des molécules et de leurs récepteurs Fc sur la surface des granulocytes va activer les cellules pour libérer rapidement les médiateurs préformés de ses granules.
Mode de fonctionnement des récepteurs
Les récepteurs Fc sont des molécules membranaires exprimées par plusieurs cellules hématopoïétiques qui reconnaissent la région Fc de plusieurs classes et sous-classes d'immunoglobulines (Ig). On distingue le FcR pour IgG (FcγRI / CD64, FcγRII / CD32 et FcγRIII / CD16), IgE (FcϵRI), IgA (FcαRI / CD89), IgM (FcμR) et IgA / IgM (Fcα / μR). Plusieurs autres récepteurs exprimés sur différents types de cellules se lient également aux molécules d'Ig : FcR néonatal pour IgG (FcRn) sur l'épithélium intestinal, le placenta et l'endothélium, FcϵR de faible affinité (FcϵRII / CD23) sur les cellules B et les macrophages, et récepteur d'Ig polymère (pIgR) sur l'épithélium muqueux La fonction des anticorps dépend d'une part de leur capacité à reconnaître les épitopes antigéniques et, d'autre part, de leur flexibilité dynamique et de leur capacité à interagir avec leurs FcR apparentés. L'engagement des FcR exprimés par les leucocytes déclenche un certain nombre de fonctions proinflammatoires, anti-inflammatoires et immunomodulatrices dans les réponses immunitaires adaptatives de l'hôte conduisant à une protection mais parfois aussi à une maladie.
Des immunoglobulines G
Activation

Les récepteurs Fc gamma générent des signaux à l'intérieur de leurs cellules via un important motif d'activation appelé motif d’activation des récepteurs immuns basé sur la tyrosine (ITAM). Un ITAM est une séquence spécifique d'acides aminés (YXXL) apparaissant deux fois de façon rapprochée dans la partie intracellulaire d'un récepteur. Lorsque des groupes phosphate sont ajoutés à des résidus de tyrosine (Y) de l'ITAM par des enzymes appelées tyrosine kinases, une cascade de signalisation est générée à l'intérieur de la cellule. Cette réaction de phosphorylation suit généralement l'interaction d'un récepteur Fc avec son ligand. Un ITAM est présent dans la queue intracellulaire de FcyRIIA, et sa phosphorylation induit la phagocytose des macrophages. FcyRI et FcyRIIIA n'ont pas d'ITAM mais peuvent transmettre une activation du signal à leurs phagocytes par l'intermédiaire d'une autre protéine. Cette protéine adaptatrice est appelée sous-unité Fcy et, comme FcyRIIA, contient les deux séquences YXXL qui sont caractéristiques d'un ITAM.
Inhibition
L'activation des récepteurs immunitaires porteurs d'ITAM peut être rétrocontrôlée par des FcR inhibiteurs porteurs d'ITIM tels que le FcγRIIB. Le motif ITIM est défini par une seule séquence [I / V / L / S] xYxx [L / V]). Cependant, l'inhibition de l'activation cellulaire par ce motif nécessite une co-ligature entre les récepteurs activateurs inhibiteurs et hétérologues par des complexes immuns favorisant le recrutement des inositol phosphatases (SHIP-1 et SHIP-2). Un autre mécanisme inhibiteur a été récemment identifié qui implique ITAM lui-même. En effet, à la suite d'interactions de ligands à faible avidité, les FcR portant ITAM induisent un signal inhibiteur soutenu sans co-ligature avec des récepteurs hétérologues. Ce mécanisme est impliqué dans le maintien de l'homéostasie immunitaire
FcγRIIB1 et FcγRIIB2 présentent des séquences ITIM et sont des inhibiteurs des récepteurs Fc ; ils n'induisent pas de phagocytose. L'action inhibitrice de ces récepteurs est contrôlée par des enzymes qui éliminent les groupes de phosphates à partir de résidus de tyrosine; les phosphatases PTPN6 (en) et INPP5D (en) inhibent la signalisation par les récepteurs Fcγ. La liaison du ligand à FcγRIIB conduit à la phosphorylation de la tyrosine du motif ITAM . Cette modification génère un site de liaison de la phosphatase, un domaine de reconnaissance SH2. L'arrêt du signal d'activation de l'ITAM est causé par l'inhibition des protéines tyrosine kinases de la famille Src, et par hydrolyse de la membrane PIP3, qui interrompt la signalisation aval par les récepteurs d'activation, telles que l'activation par FcγRs, TCR, BCR et les récepteurs de cytokines (par exemple, c-Kit).
La signalisation négative par FcγRIIB est surtout importante pour la régulation des lymphocytes B activés. La signalisation positive des lymphocytes B est initiée par la liaison de l'antigène étranger à la surface des immunoglobulines. Le même anticorps spécifique à l'antigène est sécrété et il peut le supprimer or déclencher un signal négatif. Ce signal négatif est fourni par FcγRIIB : ((en)) Experiments using B cell deletion mutants and dominant-negative enzymes⇔des expériences utilisant des mutations de cellules et des enzymes à dominant négatif ont établi le rôle important dans la signalisation négative du domaine SH2 contenant de l'inositol 5-phosphatase (SHIP). La signalisation négative par SHIP semble inhiber la voie de signalisation Ras par le biais d'une concurrence entre le domaine SH2 et les Grb2 et Shc ; elle peut impliquer la consommation de médiateurs lipides intracellulaires qui agissent comme activateurs d'enzymes allostériques ou qui favorisent l'entrée du Ca2+ extracellulaire.
Des immunoglobulines A et M
Un autre FcR qui joue un rôle essentiel dans la transcytose des cellules épithéliales des IgA dimères, mais aussi des IgM pentamériques (notamment lors des déficits en IgA), est appelé le récepteur polymérique des immunoglobulines (pIgR). Le récepteur polymérique des immunoglobulines est internalisé avec ses ligands par endocytose et transcyté de la membrane basolatérale vers le côté apical de la cellule épithéliale . Le rôle central de ce récepteur est de générer des IgA sécrétoires (formées de dimères d'IgA liés au domaine extracellulaire du pIgR, également connu sous le nom de composant sécrétoire) dans les sécrétions exocrines pour établir une symbiose hôte-microbiote et assurer la médiation de la protection des surfaces muqueuses contre les agents pathogènes. Le Fcα / μR, le récepteur Fc des IgA et IgM, peut jouer un rôle dans l'immunité systémique et muqueuse. Il a été démontré qu'aucun des lymphocytes B, des lymphocytes T, des monocytes / macrophages ou des cellules NK dans les échantillons de sang humain n'exprimait ce récepteur indépendamment de l'âge, de l'origine ethnique ou du sexe . Son expression est limitée aux cellules B du centre germinal, aux cellules dendritiques folliculaires (ganglion lymphatique) et aux cellules des amygdales. Bien que la fonction exacte du Fcα / μR ne soit pas entièrement clarifiée, elle peut jouer un rôle important dans la présentation de l'antigène et la sélection des cellules B dans les réponses du centre germinal
Activation cellulaire
Sur les phagocytes
Lorsque des molécules d'IgG, spécifiques à un antigène ou à des composants membranaires donnés, se lient à l'agent pathogène dans leurs régions Fab (fragment antigen binding région), leurs régions Fc pointent vers l'extérieur, à portée immédiate des phagocytes. Les phagocytes lient ces régions Fc régions avec leurs récepteurs Fc. De nombreuses interactions de faible affinité se produisent entre les récepteurs et les anticorps qui travaillent ensemble pour se lier fermement aux microbes enveloppés d'anticorps. La faible affinité individuelle empêche les récepteurs Fc de se lier aux anticorps en l'absence d'antigène, et, par conséquent, réduit la probabilité de l'activation des cellules immunitaires en l'absence d'infection. Cela empêche également l'agglutination des phagocytes par des anticorps lorsqu'il n'y a pas d'antigène. Après qu'un agent pathogène a été lié, les interactions entre la région Fc de l'anticorps et les récepteurs Fc des phagocytes provoquent l'initiation de le phagocytose. L'agent pathogène est englouti par les phagocytes par un processus actif impliquant la liaison et la libération du complexe région Fc/récepteur Fc, jusqu'à ce que la membrane cellulaire de la phagocyte recouvre entièrement l'agent pathogène.
Sur les lymphocytes NK
Les récepteurs Fc sur les cellules NK qui reconnaissent les IgG liées à la membrane d'une cellule-cible infectée par un agent pathogène sont appelées CD16 ou FcγRIII. L'activation de FcγRIII par les IgG provoque la libération de cytokines telles que l'IFN-γ qui envoie des signaux à d'autres cellules immunitaires et médiateurs cytotoxiques comme la perforine et le granzyme, lesquelles entrent dans la cellule cible et déclenchent la mort cellulaire par apoptose. Ce processus est connu sous le nom de Cytotoxicité à médiation cellulaire dépendante des anticorps (ADCC). Les récepteurs FcγRIII présents sur les cellules NK peuvent également être associés avec des IgG monomères (c'est-à-dire des IgG non liées à des antigènes). Lorsque cela se produit, le récepteur Fc inhibe l'activité des lymphocytes NK.
Sur les mastocytes

Les anticorps IgE se lient aux antigènes des allergènes. Ces allergènes liés à des molécules d'IgE interagissent avec les récepteurs Fcε à la surface des mastocytes. L'activation des mastocytes après l'engagement de FcεRI provoque un processus appelé la dégranulation, dans lequel les mastocytes libèrent des molécules préformées à partir de leurs granules cytoplasmiques. Celles-ci sont un mélange de composés incluant histamine, protéoglycanes, et protéases à sérine Les mastocytes activés synthétisent et sécrétent aussi des médiateurs lipidiques (tels que prostaglandine, leucotriène, et platelet-activating factor) et des cytokines (interleukine 1, interleukine 3, interleukine 4, interleukine 5, interleukine 6, interleukine 13, facteur de nécrose tumorale-alpha, GM-CSF, et plusieurs chimiokines.. Ces médiateurs contribuent à l'inflammation en attirant d'autres leucocytes.
Sur les éosinophiles
Les gros parasites comme les helminthes (vers) Schistosoma mansoni sont trop grands pour être ingérés par les phagocytes. Ils ont aussi une structure externe appelé tégument qui est résistant à l'attaque par des substances libérées par les macrophages et les mastocytes. Cependant, ces parasites peuvent être recouverts par des IgE et reconnus par FcεRI sur la surface des éosinophiles. Les éosinophiles activés libérent des médiateurs préformés comme la protéine basique majeure, et des enzymes telles que la peroxydase, contre qui les helminthes ne sont pas résistants. L'interaction de la FcεRI récepteur avec la portion Fc des helminthes lié IgE entraîne la libération par les éosinophiles de ces molécules dans un mécanisme similaire à celui des cellules NK au cours de l'ADCC.
Différents types de récepteurs


Les FcR sont divisés en type I et type II sur la base de l'état de conformation du domaine Ig Fc qui interagit avec le récepteur
- Les récepteurs Fc de type I interagissent avec la conformation Ig Fc «ouverte», mais pas «fermée». Ces récepteurs comprennent FcγRI, FcγRII, FcγRIII, FcεRI, FcαRI, FcμR et Fcα / μR . En revanche, les FcR de type II se lient préférentiellement aux domaines Ig Fc en conformation «fermée». Parmi ceux-ci figurent des récepteurs de lectine de type C tels que FcεRII (CD23) et le récepteur DC-SIGN.
- Pour les récepteurs Fc de type II, la glycosylation du domaine Fc induit un changement de conformation qui obstrue le site de liaison pour les récepteurs Fc de type I qui se trouve près de la région charnière de l'immuglobuline (conformation ouverte) et révèle un site de liaison à l'interface du domaine CH2-CH3 (conformation fermée ). Ces récepteurs se lient aux anticorps dans une stœchiométrie à deux récepteurs contre un qui peut influencer l'initiation du signal . DC-SIGN et SIGN-R1, par exemple, se lient aux IgA sécrétoires et jouent un rôle dans les cellules dendritiques induisant une tolérance médiée par interleukine 10 (connue pour son action inhibitrice sur la réaction inflammatoire) et Threg . La signalisation à travers ces récepteurs, cependant, n'est pas encore documentée par rapport aux FcR de type I, à l'exception du CD23. La modification du CD23 sur les cellules B active l'AMPc et le flux de calcium intracellulaire qui est associé à l'activation du SFK Fyn et de la voie PI3K . Ces résultats sont en accord avec les données récentes sur les FcR de type I , et indiquent que Fyn joue également un rôle d'activation dans les cellules B via les récepteurs Fc de type II.
Classes
Il existe plusieurs types de récepteurs Fc (en abrégé FcR), qui sont classés en fonction du type d'anticorps qu'ils reconnaissent. La lettre latine utilisée pour identifier un type d'anticorps est remplacée par la lettre grecque correspondante, après " Fc " pour former le nom du récepteur. Par exemple, ceux qui se lient le plus communément à la classe des anticorps IgG, sont appelés récepteurs Fc-gamma (FcyR) ; ceux qui se lient à IgA sont appelés récepteurs Fc-alpha (FcαR) et ceux qui se lient à IgE sont appelés récepteurs Fc-epsilon (FcεR).
On distingue aussi les classes de FcR en fonction des cellules qui les expriment (macrophages, granulocytes, lymphocytes NK, lymphocytes B, cellules T et B) et des propriétés de signalisation de chaque récepteur.
Récepteurs Fc-gamma

L'ensemble des récepteurs Fcγ (FcγR) appartiennent à la superfamille des immunoglobulines et les plus importants sont les récepteurs Fc induisant la phagocytose de microbes opsonisés (marqués). Cette famille comprend plusieurs membres, FcγRI (CD64), FcγRIIA (CD32), FcγRIIB (CD32), FcγRIIIA (CD16a), FcγRIIIB (CD16b), qui diffèrent dans leurs affinités anticorps en raison de leur différente structure moléculaire. Par exemple, FcyRI se lie à IgG plus fortement que FcγRII ou FcγRIII. FcγRI a aussi une partie extracellulaire composé de trois domaines de type immunoglobuline (Ig), soit un de plus que FcγRII ou FcγRIII. Cette propriété permet à FcγRI de se lier à une molécule isolée d'IgG (ou monomère), mais tous les récepteurs Fcγ doivent se lier à plusieurs molécules d'IgG au sein d'un complexe immunitaire pour être activés.
Les récepteurs Fc-gamma se distinguent par leur affinité pour les IgG et de la même manière que les différentes sous-classes d'IgG ont des affinités pour chacun des récepteurs Fc gamma. Ces interactions sont de plus régulées par le glycane (oligosaccharides) présent à la position CH2-84.4 des IgG. Par exemple, en créant un encombrement stérique, la fucose contenant des glycanes CH2-84.4 réduit l'affinité des IgG pour FcγRIIIA. En revanche, les glycanes G0, qui manquent de galactose et se terminent par des groupements GlcNAc, ont une affinité accrue pour FcγRIIIA.
Un autre FcR est exprimé sur plusieurs types de cellules et possède une structure similaire à celle du Complexe majeur d'histocompatibilité de classe I. Ce récepteur se lie également aux IgG et est impliqué dans la préservation de cet anticorps. Toutefois, comme ce récepteur Fc est également impliqué dans le transfert des IgG de la mère via le placenta à son fœtus ou via le lait au nourrisson qu'elle allaite, il est appelé récepteur Fc néonatal (FcRn).
Le récepteur Fcγ couvre une région chevauchant partiellement le domaine de liaison du facteur de complément C1q. Pour toutes les interactions du récepteur Fcγ, l'étirement des acides aminés comprenant l'extrémité N-terminale des domaines et des brins CH2 adjacents dans le repli d'immunoglobuline tridimensionnelle est important pour la liaison. En général, cela englobe les acides aminés 234-239, 265-269, 297-299 et 327-330 Cependant, chacune des sous-classes d'IgG a un profil de liaison unique à chaque récepteur Fcγ, et leurs profils d'expression sont très variables entre les différentes cellules immunitaires d'origine myéloïde et NK . Une distinction majeure peut être faite entre les IgG1 / IgG3 qui interagissent efficacement avec la plupart des récepteur Fcγ et les IgG2 / IgG4, qui montrent une affinité réduite pour un certain nombre de récepteur Fcγ. De plus, l'IgG3 monomère se lie plus efficacement que l'IgG1 monomère à FcγRIIa, FcγRIIIa et FcγRIIIb, et l'efficacité de liaison de l'IgG3 à tous les récepteur néonatals Fcγ s dépasse celle de l'IgG1. Les déterminants structurels responsables des différences entre IgG1 et IgG3 sont encore inconnus.
Les IgG peuvent être divisées en quatre sous-classes, nommées, par ordre décroissant d'abondance IgG1, IgG2, IgG3 et IgG4
Récepteur Fc-gamma I (FcγRI)
Bien que FcγRI soit souvent considéré comme une seule entité, FcγRI se compose de trois gènes homologues sur le bras court du chromosome un , et plusieurs variantes d'épissage alternatives ont été décrites . Cependant, seul le FcγRIa constitué de trois domaines extracellulaires a été étudiée en détail. Le gène codant le variant potentiel de FcγRIb, consiste potentiellement en un récepteur presque identique avec seulement les deux domaines extracellulaires Ig N-terminaux, mais conservant la partie cytoplasmique intracellulaire, tandis que FcγRIc n'aurait pas non plus de partie cytoplasmique et de région transmembranaire, et serait, par conséquent, il devrait être prévu de représenter une forme sécrétée. Le récepteur FcγRIa se lie à toutes les sous-classes d'IgG humaines à l'exception d'IgG2, et contrairement aux autres FcγR, contient son troisième domaine d'immunoglobuline membranaire proximal unique qui est également responsable de sa plus grande affinité pour les IgG .
Récepteur Fc-gamma IIa (FcγRIIa)
Le FcγRIIa est le FcγR le plus largement exprimé sur les cellules myéloïdes et a été décrit comme le seul FcγR avec une liaison significative aux IgG2 La liaison est plus efficace pour la variante 131H («faible réponse», LR) que la variante 131R («haute réponse», HR) . L'affinité de liaison varie selon les sous-classes comme suit: IgG3> IgG1> IgG4 = IgG2.
Récepteur Fc-gamma IIB (FcγRIIB)
Il existe trois isomères connus de FcγRIIB chez la souris et chez l'homme. FcγRIIB1 et FcγRIIB2, sont des formes intermenbranaires de surface. FcγRIIB3 est une forme soluble, relativement peu explorée in vivo.
La forme la plus répandue, FcγRIIB1, est aussi la seule à être exprimée par les lymphocytes B. Lorsqu'il est réticulé avec les récepteur des cellules B, FcγRIIB augmente de le seuil d'activation des lymphocytes, et diminue leur production d'anticorps. Cette fonction est due à la présence de motifs ITIM (en), qui recrute une enzyme SHIP (en) pour bloquer l'action de la PI-3-kinase. FcγRIIB1 est exprimé dans une moindre mesure par les monocytes.
Lorsqu'il est exprimé par les cellules dendritiques, les macrophages, les neutrophiles activés, les mastocytes et les basophiles, FcyRIIB a un rôle d'inhibiteur des fonctions d'activation des FcyR, telles que la phagocytose et la libération de cytokines proinflammatoires.
Récepteur Fc-gamma IIb/IIc (FcγRIIb/IIc)
Le domaine extracellulaire du FcγRIIb inhibiteur est identique au FcγRIIc activateur qui est exprimé chez ~ 11% des individus[pas clair][citation nécessaire]. La liaison au récepteur inhibiteur FcyRIIb ou IIc est faible pour toutes les sous-classes, préférant généralement IgG3 = IgG1 = IgG4> IgG2. Fait intéressant, les constantes de dissociation pour la liaison des IgG1 et IgG3 monomères sont similaires, mais les complexes immuns des IgG3 semblent se lier plus efficacement par rapport aux IgG1 . La liaison à la plupart des FcRns activateurs est plus faible pour l'IgG4 que pour l'IgG1, mais ce n'est pas le cas pour le récepteur inhibiteur FcγRIIb. Cet équilibre modifié entre la liaison aux récepteurs activateurs par rapport aux récepteurs inhibiteurs peut être une caractéristique importante de l'IgG4 qui contribue à sa faible capacité proinflammatoire.
Récepteur Fc-gamma IIIa (FcγRIIIa)
Il existe deux variantes allotypiques de FcγRIIIa: F158 et V158. La variante V158 a une plus grande affinité pour toutes les sous-classes, et pour l'IgG3, l'efficacité de liaison approche celle de FcyRIa, avec des affinités générales suivant IgG3> IgG1>> IgG4> IgG2. En plus de changer les acides aminés 233–236 d'IgG1 en équivalents d'IgG2, A327G (Alanine présent dans IgG1 et IgG3, Glycine dans IgG2 et IgG4) entraîne également une diminution de la liaison .
Récepteur Fc-gamma IIIb (FcγRIIIb)
Il existe des variations allotypiques fonctionnelles des FcγRIIIb des neutrophiles , appelés antigène 1 des neutrophiles humains (NA1 / HNA1a) et (NA2 / HNA1b) (162). La forme FcγRIIIb-NA1 est capable de mieux ingérer des particules opsonisées IgG1 ou IgG3 que RIIIb-NA2 . Le FcγRIIIb se lie généralement aux IgG1 et IgG3 mais pas aux IgG2 et IgG4, les IgG3 se liant mieux que les IgG1 . Une structure cristalline du complexe d'IgG1 Fc avec FcγRIIIb révèle que les acides aminés 234-238 sont d'importants résidus de contact, et la variation spécifique à la sous-classe dans ce domaine peut à nouveau expliquer le manque de liaison des IgG2 et IgG4 à ce récepteur
Récepteur Fc-gamma néonatal (FcγRn)
Dans les années 1960, l'existence d'un récepteur responsable de la demi-vie inhabituellement longue des IgG (3 semaines) et d'un transport efficace de ces IgG de la mère à l'enfant a d'abord été proposée Cela a été confirmé par la suite par divers groupes et finalement cloné et identifié comme le FcRn néonatal.
Le FcRn est étonnamment similaire aux molécules du CMH de classe I. Comme le MHC classe I et d'autres molécules de type MCH classe I, le FcRn est coexprimé avec la microglobuline non glycosylée β2 12 kD, codée sur le chromosome 15. La chaîne α du FcRn humain, une chaîne polypeptidique de 45 kD, est codé sur le chromosome 19 à un locus hébergeant divers autres récepteurs immunitaires (par exemple, KIR, LAIR-1, CD89, CEACAM). Le FcRn humain n'a qu'un seul site de glycosylation potentiel (N102). Il est situé sur la face opposée au site de liaison aux IgG et est également partagé avec celui de toutes les séquences connues de FcRn . FcRn ne lie pas son ligand à pH physiologique (7,4). Ce n'est que dans l'environnement acide des vacuoles endocytaires (pH ≤ 6,5) Les résidus d'histidine dans la partie Fc des IgG (interface CH2-CH3) sont essentiels pour la liaison de haute affinité aux résidus dans la chaîne α β2M et FcRn H435 se trouve au cœur de cette interface. Par conséquent, l'IgG3 a une demi-vie normale de 3 semaines et est transportée efficacement à travers le placenta chez les individus contenant des allotypes d'IgG3 contenant H435 .
Le fait que le FcRn protège les IgG de la dégradation a été confirmé par des modèles murins : la demi-vie des IgG est diminuée chez les souris déficientes en FcRn ou en β2-microglobuline. Le recyclage induit par le FcRn permet de conserver quatre fois plus d'IgG que ce qui est produit. Bien qu'il ait été initialement proposé que l'expression de FcRn sur les cellules endothéliales soit responsable du recyclage des IgG , des études ultérieures ont montré que la forte expression de FcRn sur les cellules myéloïdes contribue également à l'extension de la demi-vie chez la souris. De même, la surexpression de FcRn chez les animaux transgéniques entraîne des taux sériques d'IgG plus élevés .
Cependant, le FcRn commence sa fonction tôt dans la vie par le transport des IgG - et donc l'immunité humorale - à travers le placenta de la mère à l'enfant et chez les rongeurs également après la naissance par le transport du lait maternel dans l'intestin des nouveau-nés allaités. Chez le rat, cette expression de FcRn est régulée à la baisse dans l'intestin grêle, ce qui est en corrélation avec la dégradation des IgG dans ces cellules.
Rôle des récepteurs Fc-gamma néonatals chez l'adulte
Chez l'adulte, le FcRn est exprimé sur de nombreuses cellules épithéliales et continue de fonctionner dans le transport des IgG à travers les barrières épithéliales d'expression du FcRn . Le FcRn est capable (chez toutes les espèces) de transiter de façon bidirectionnelle à travers les cellules polarisées (épithéliales et endothéliales), mais la direction du transport net dépend du tissu .Les complexes d'immunoglobulines G ou d'IgG-antigènes sont transportés à travers les surfaces muqueuses, telles que la cavité intestinale ou l'épithélium respiratoire, et fonctionnaient ainsi dans la surveillance immunitaire . Avec ce rôle dans l'immunité muqueuse, il complète sIgA dans la fonction immunorégulatrice .
Comme les IgG peuvent transporter des protéines fonctionnelles à travers les barrières épithéliales, cela offre de nouvelles possibilités au FcRn en tant que récepteur endogène pour transporter les protéines ou les antigènes vaccinaux à travers des surfaces épithéliales.
Sur les cellules muqueuses, le FcRn s'est révélé transporter les IgG et être impliqué dans l'échantillonnage des antigènes ), et son expression sur les cellules phagocytaires s'est récemment révélée améliorer la capacité de phagocytose des particules opsonisées par les IgG . Sur les cellules présentant l'antigène, cette ingestion de complexes IgG peut conduire à une présentation améliorée . Semblable aux réponses de phagocytose, la présentation améliorée nécessite probablement la détection externe et l'activation cellulaire via le FcγR et les récepteurs de reconnaissance de formes, reconnaissant le complexe d'IgG-antigène au FcRn à faible pH . Ainsi, les activités d'immunoglobulines, y compris la demi-vie prolongée, le transport et l'échantillonnage des antigènes, semblent être orchestrées via un seul récepteur, le FcRn de type I du CMH. En revanche, d'autres fonctions effectrices des IgG, telles que la phagocytose et la présentation de l'antigène semblent être médiées à la fois par le FcRn et les FcγR classiques.
Récepteurs Fc- alpha
Un seul récepteur Fc appartient au sous-groupe Fcα , il s'agit de FcαRI (ou CD89). FcαRI se trouve sur la surface des neutrophiles, des éosinophiles, monocytes, de certains macrophages (y compris les cellules de Kupffer), et de quelques cellules dendritiques. Il est composé de deux domaines extracellulaire de type Ig, et fait partie à la fois de la superfamille des immunoglobulines et de la famille des récepteurs de reconnaissance immunitaire multi-chaîne (MIRR). Il signale en s'associant avec deux FcRy de signalisation des chaînes. Un autre récepteur peut également se lier aux IgA, même s'il a une plus grande affinité pour un autre anticorps, l'immunoglobuline M. Ce récepteur est appelé récepteur Fc-alpha/mu (Fcα/µR) et est un type I de protéine transmembranaire. Avec un domaine de type Ig dans sa partie extracellulaire, ce récepteur Fc est également un membre de la superfamille des immunoglobulines.
Récepteurs Fc-epsilon
Deux types de FcεR sont connus :
- Le récepteur FcεRI de forte affinité est un membre de la superfamille des immunoglobulines (il a deux domaines de type Ig). On trouve FcεRI sur les cellules de Langerhans épidermiques, les éosinophiles, les mastocytes et les basophiles En raison de sa distribution cellulaire, ce récepteur joue un rôle majeur dans le contrôle des réactions allergiques. FcεRI est également exprimé sur les cellules présentatrices de l'antigène, et contrôle la production d'importants médiateurs immunitaires appelées cytokines qui favorisent l'inflammation.
- le récepteur de faible affinité FcεRII (CD23) est une lectine de type C. FcεRII a de multiples fonctions qu'il soit lié à la membrane ou comme récepteur soluble ; il contrôle la croissance et la différenciation des cellules B et bloque la liaison IgE des éosinophiles, monocytes et des basophiles.
TRIM21
Ce récepteur Fc occupe une place à part. L'interaction TRIM21 (tripartite motif containing 21)/IgG a été mise en évidence en 1999, et sa fonction de récepteur admise en 2008. Il a deux fonctions.
- La première, antivirale, s'effectue sur des virus liés à des anticorps, mais ayant réussi à entrer dans le cytoplasme de la cellule attaquée. Le mécanisme mis en œuvre, appelé neutralisation cellulaire dépendante des anticorps (ADIN) désactive et détruit le virus via une Ubiquitine-ligase (E3) et par recrutement de la machinerie protéasomique.
- La seconde fonction est une activité de signalisation immunitaire.
Ses deux particularités sont son large spectre d'action (il s'adresse aux Fc des IgA, TgM et IgG) et le fait d'être exprimé principalement par des cellules de lignées histogéniques (c'est-à-dire à l'origine de la fabrication de tissus), tandis que l'expression des FcyR classiques est principalement limitée aux cellules hématopoïétiques (produisant des cellules sanguines).
Tableau récapitulatif
| Nom du récepteur | Principal ligand anticorps | Affinité pour le ligand | Cellules concernées | Fonctions | Effet résultant de la liaison aux anticorps | Maladies liées |
| Récepteur Fc des immunoglobulines G | ||||||
| FcγRI (CD64) | IgG1 et IgG3 | Forte (Kd ~ 10−9 M) |
Macrophages |
Activation | Phagocytose Activation cellulaire |
|
| FcγRIIA (CD32) | IgG | Faible (Kd > 10−7 M) | Macrophages Neutrophiles |
Activation
Inhibition |
Phagocytose Dégranulation (eosinophiles) |
Maladie de Kawasaki Rectocolite hémorragique Purpura thrombopénique immunologique |
| FcγRIIB (CD32b) | IgG | Faible(Kd > 10−7 M) |
Lymphocyte B |
Inhibition | Absence de phagocytose Inhibition de l'activité cellulaire |
|
| FcγRIIC (CD32c) | IgG | Faible (Kd > 10−7 M) | Macrophages Neutrophiles |
Activation | Phagocytose Inhibition de l'activité cellulaire |
Maladie de Kawasaki |
| FcγRIIIA (CD16a) | IgG | Faible (Kd > 10−6 M) |
NK cells Macrophages (certain tissues) |
Activation
Inhibition |
Induction de la cytotoxicité à médiation cellulaire dépendante des anticorps (ADCC) Induction de la libération de cytokine release par les macrophages |
|
| FcγRIIIB (CD16b) | IgG | Faible (Kd > 10−6 M) | Eosinophiles Macrophages |
Activation | Induction de la lyse microbienne | |
| Récepteur Fc des immunoglobulines E | ||||||
| FcεRI | IgE | Forte (Kd ~ 10−10 M) | Blastocytes Eosinophiles |
Degranulation Phagocytose |
||
| FcεRII (CD23) | IgE | Faible (Kd > 10−7 M) | Leucocytes B Eosinophiles |
Possible adhésion moléculaire transport d'IgE à travers l'épithelium intestinal |
||
| Récepteur Fc des immunoglobulines A | ||||||
| FcαRI (CD89) | IgA | Faible (Kd > 10−6 M) | Monocytes Macrophages |
Phagocytose Induction de la lyse microbienne |
||
| Fcα/μR | IgA et IgM | Forte pour IgM, Moyenne pour IgA | Lymphocytes B Cellules mésangiales |
Endocytose Induction de la lyse microbienne |
||
| Récepteur Fc néonatal | ||||||
| FcRn | IgG | Monocytes Macrophages |
Transfère les IgG de la mère au fœtus via le placenta Transfère les IgG de la mère à l'enfant par le lait |
|||
Récepteur Fc et maladie
Utilisation thérapeutique des récepteurs Fc
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Receptor Fc » (voir la liste des auteurs).
Notes
Références
Annexes
Articles connexes
Bibliographie
- Ben Mkaddem S, Benhamou M and Monteiro RC (2019) Understanding Fc Receptor Involvement in Inflammatory Diseases: From Mechanisms to New Therapeutic Tools. Front. Immunol. 10:811. doi: 10.3389/fimmu.2019.00811 Article sous licence CC-BY
- Vidarsson G, Dekkers G and Rispens T (2014) IgG subclasses and allotypes: from structure to effector functions. Front. Immunol. 5:520. doi: 10.3389/fimmu.2014.00520 Article sous licence CC-BY-SA