
| N° EC | EC |
|---|---|
| N° CAS | |
| Cofacteur(s) | FMN ; Fe-S |
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
| GO | AmiGO / EGO |
La NADH déshydrogénase, ou complexe I de la chaîne respiratoire, est une oxydoréductase membranaire qui catalyse la réaction :
- NADH + ubiquinone + 5 H+matriciel NAD+ + ubiquinol + 4 H+intermembranaire.

Cette enzyme est présente chez un très grand nombre d'êtres vivants, allant des procaryotes jusqu'aux humains. Elle constitue le premier point d'entrée des électrons à haut potentiel de transfert dans la respiration cellulaire et la phosphorylation oxydative dans les mitochondries et assure le transfert des électrons du NADH vers la coenzyme Q10 dissoute dans la bicouche lipidique. Chez les eucaryotes, elle est située dans la membrane mitochondriale interne. Elle jouerait également un rôle dans le déclenchement de l'apoptose ; une corrélation a pu être établie entre l'activité des mitochondries et l'apoptose lors du développement somatique de l'embryon.
Structure et fonctionnement
Le complexe I est l'enzyme la plus grande et la plus compliquée de la chaîne respiratoire. Chez les mammifères, elle est constituée de 44 chaînes polypeptidiques, dont sept sont encodées par le génome mitochondrial. Elle contient notamment un groupe prosthétique FMN et huit clusters fer-soufre dont sept sont alignés pour permettre la circulation des électrons depuis le NADH vers la coenzyme Q10. Le transfert de ces électrons d'un couple rédox dont le potentiel standard est −0,32 V vers un couple rédox dont le potentiel standard est +0,06 V libère suffisamment d'énergie pour actionner une pompe à protons expulsant quatre protons H+ de la matrice mitochondriale vers l'espace intermembranaire à travers la membrane interne de la mitochondrie.
La structure générale adopte une forme en L avec un long domaine membranaire constitué d'une soixantaine d'hélices et un domaine hydrophile périphérique où se trouvent tous les centres rédox et le site de liaison au NADH. La structure du complexe I des eucaryotes n'est pas encore bien caractérisée dans son ensemble mais celle du domaine hydrophile de la bactérie Thermus thermophilus (en) a pu être établie (PDB 2FUG) ainsi que celle des domaines membranaires d'E. coli (PDB 3RKO) et de T. thermophilus (PDB 4HE8). La structure complète d'une NADH déshydrogénase de T. thermophilus a été publiée pour la première fois en (PDB 4HEA).
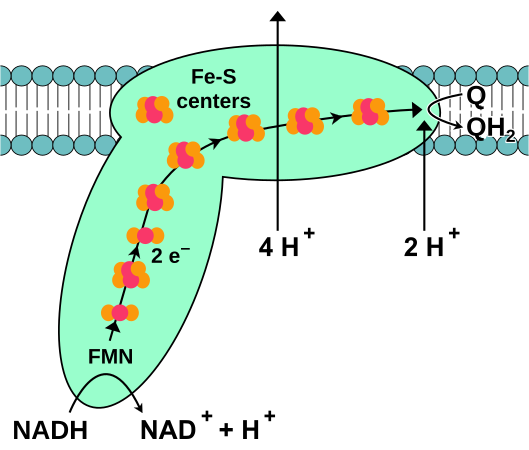
(en) Schéma de fonctionnement de la NADH déshydrogénase montrant sa forme générale constituée d'un domaine hydrophile (en bas) baignant dans la matrice mitochondriale et jalonné par sept clusters fer-soufre alignés conduisant les électrons issus du NADH matriciel via un groupe prosthétique FMN jusqu'au domaine hydrophobe (en haut) inclus dans la membrane mitochondriale interne et dans laquelle la coenzyme Q10 est réduite en ubiquinol Q10H2. Quatre protons H+ sont expulsés vers l'espace intermembranaire mitochondrial au cours de cette réaction, ce qui contribue à générer un gradient de concentration de protons à travers la membrane mitochondriale interne.

Inhibiteurs
La roténone est l'inhibiteur le plus connu du complexe I. Elle est couramment utilisé comme pesticide et comme insecticide. Avec les isoflavonoïdes (en), les roténoïdes sont présents chez plusieurs genres de plantes tropicales telles que Antonia (Loganiaceae), Derris et Lonchocarpus (Faboideae, Fabaceae). Tout comme la piéricidine A (en), la roténone se lie au site de liaison à l'ubiquinone du complexe I.
L'adénosine diphosphate ribose (ADPR) est également un inhibiteur de la NADH déshydrogénase en se liant au site de liaison nucléotidique, ce qui bloque la liaison du NADH.
La famille des acétogénines forme les inhibiteurs les plus puissants du complexe I. Ces molécules se lient à la sous-unité ND2, ce qui tend à montrer que celle-ci serait essentielle à la liaison de l'ubiquinone. L'une de ces molécules, la rolliniastatine-2, est le premier inhibiteur du complexe I qui ne se lie pas au même site que la roténone.
La metformine, un antidiabétique, est également un inhibiteur partiel du complexe I, ce qui semble jouer un rôle essentiel dans son mode d'action.