
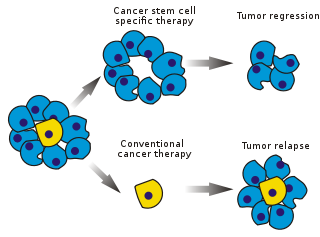
Les cellules souches cancéreuses (CSCs) sont des cellules cancéreuses (présentes dans les tumeurs dites « solides » ou les cancers hématologiques) qui possèdent des caractéristiques associées aux cellules souches normales, notamment la capacité de donner naissance aux différentes populations de cellules présentes dans une tumeur particulière. Les CSCs sont donc tumorigènes (formant des tumeurs), peut-être à la différence d'autres cellules cancéreuses non tumorigènes. Les CSCs peuvent générer des tumeurs grâce aux processus d'auto-renouvellement et de différenciation des cellules souches vers l’ensemble des composants cellulaires formant la tumeur. On suppose que ces cellules persistent dans les tumeurs comme une population distincte et provoquent des rechutes et des métastases en donnant naissance à de nouvelles tumeurs. Par conséquent, le développement de thérapies spécifiques ciblées sur les CSCs permet d'espérer une amélioration de la survie et de la qualité de vie de patients atteints de cancer, en particulier pour les patients atteints de maladies métastatiques.
Les traitements existants contre le cancer ont été principalement développés sur la base de modèles animaux, où les thérapies capables de favoriser la réduction des tumeurs ont été jugées efficaces. Cependant, les animaux ne reproduisent pas de manière fidèle la maladie humaine. En particulier, chez les souris, dont la durée de vie ne dépasse pas deux ans, la rechute des tumeurs est difficile à étudier.
L'efficacité des traitements anticancéreux est, dans les premiers stades des tests, correspond souvent à la fraction éliminée de cellules de la masse tumorale (fractional kill). Les CSCs ne représentant qu’une faible proportion de la masse tumorale, les traitements anticancéreux n’ont pas nécessairement d’effet spécifique sur elles. La théorie suggère que les chimiothérapies conventionnelles tuent les cellules différenciées ou en cours de différenciation, qui forment la masse de la tumeur mais qui ne génèrent pas de nouvelles cellules. Une population de CSCs, ayant donné naissance à la tumeur, pourrait rester intacte et provoquer une rechute.
Les CSCs ont été identifiées pour la première fois par John Dick dans la leucémie aiguë myéloïde à la fin des années 1990. Depuis le début des années 2000, elles font l'objet de recherches intensives en oncologie. Le terme de CSC a été inventé par les biologistes Tannishtha Reya, Sean J. Morrison, Michael F. Clarke et Irving Weissman dans un article publié dans Nature en 2001.
Modèle de propagation tumorale
Dans différents sous-types de tumeurs, les cellules de la population tumorale présentent une hétérogénéité fonctionnelle et les tumeurs sont formées de cellules ayant des capacités de prolifération et de différenciation différentes. Cette hétérogénéité fonctionnelle des cellules cancéreuses a conduit à la création de multiples modèles de propagation pour tenir compte de l'hétérogénéité et des différences de capacité de régénération des tumeurs : le modèle de cellule souche cancéreuse (CSC) et le modèle stochastique. Cependant, certaines études maintiennent que cette différence est artificielle, puisque les deux processus agissent de manière complémentaire en ce qui concerne les populations tumorales réelles.


Modèle cellule souche cancéreuse
Le modèle de CSC, également connu sous le nom de modèle hiérarchique, propose que les tumeurs soient organisées de manière hiérarchique (les CSCs se trouvant au sommet (Fig. 3)). Dans la population des tumeurs, il existe des CSCs qui sont des cellules tumorigènes et qui sont biologiquement distinctes des autres sous-populations. Elles présentent deux caractéristiques déterminantes : leur capacité à s'auto-renouveler indéfiniment et leur capacité à se différencier en une descendance non tumorigène mais qui contribue quand même à la croissance de la tumeur. Ce modèle suggère que seules certaines sous-populations de CSCs ont la capacité d’induire la progression tumorale, ce qui signifie qu'il existe des caractéristiques spécifiques (intrinsèques) qui peuvent être identifiées et ensuite ciblées pour détruire une tumeur à long terme sans qu'il soit nécessaire de combattre la tumeur dans son ensemble.
Modèle stochastique
Selon le "modèle stochastique" (ou "modèle d'évolution clonale"), chaque cellule cancéreuse d'une tumeur pourrait acquérir la capacité de s'auto-renouveler et de se différencier en fonction des lignées nombreuses et hétérogènes de cellules cancéreuses qui compromettent une tumeur. Ainsi, pour qu'une cellule devienne cancéreuse, elle doit subir un nombre important d'altérations de sa séquence d'ADN. Ce modèle cellulaire suggère que ces mutations pourraient se produire dans n'importe quelle cellule du corps et provoquer un cancer. Essentiellement, cette théorie propose que toutes les cellules possèdent la capacité d'être tumorigènes, rendant toutes les cellules tumorales équipotentes avec la capacité de s'auto-renouveler ou de se différencier, conduisant à l'hétérogénéité de la tumeur tandis que d'autres peuvent se différencier en cellules non-CSCs. Le potentiel de la cellule peut être influencé par des facteurs génétiques ou épigénétiques (mutations), entraînant une diversité phénotypique des cellules tumorigènes et non-tumorigènes qui composent la tumeur.
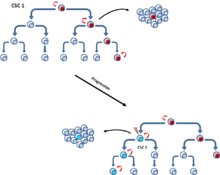
Ces mutations pourraient s'accumuler progressivement et améliorer la résistance et l'aptitude des cellules qui leur permettent de concurrencer d'autres cellules tumorales, mieux connu sous le nom de modèle d'évolution somatique. Le modèle d'évolution clonale, qui se produit à la fois dans le modèle CSC et le modèle stochastique, postule que les cellules tumorales mutantes, ayant un avantage de croissance, surpassent les autres. Les cellules de la population dominante ont un potentiel similaire pour initier la croissance d'une tumeur. (Fig. 4).

Ces deux modèles ne sont pas mutuellement exclusifs, car les CSCs elles-mêmes subissent une évolution clonale. Ainsi, d’autres CSCs peuvent émerger si une mutation leur confère des propriétés plus agressives (Fig. 5).
Relation modèle CSC et modèle stochastique
Une étude réalisée en 2014 soutient que le fossé entre ces deux modèles controversés peut être comblé en fournissant une explication alternative sur l'hétérogénéité des tumeurs. Les chercheurs ont démontré un modèle qui inclut à la fois des aspects du modèle stochastique et du modèle CSC. Ils ont examiné la plasticité des cellules souches cancéreuses, dans laquelle les cellules souches cancéreuses peuvent faire la transition entre les cellules souches non cancéreuses (non-CSCs) et les CSCs in situ, ce qui soutient un modèle plus stochastique. Toutefois, l'existence de populations biologiquement distinctes de non-CSC et de CSC soutient un modèle plus CSC, ce qui laisse entendre que les deux modèles peuvent jouer un rôle vital dans l'hétérogénéité des tumeurs.
Modèle immunologique CSC
Ce modèle suggère que les propriétés immunologiques peuvent être importantes pour comprendre la tumorigenèse et l'hétérogénéité. Ainsi, les CSCs peuvent être très rares dans certaines tumeurs, mais certains chercheurs ont découvert qu'une grande proportion de cellules tumorales peuvent initier des tumeurs si elles sont transplantées dans des souris immunodéprimées. Ces données remettent en question la pertinence d’une population rare de CSCs. Cependant, les cellules souches et les CSCs possèdent des propriétés immunologiques uniques qui les rendent plus résistantes à l'immunosurveillance. Alors, seules les CSCs peuvent être capables d'ensemencer des tumeurs chez des patients faisant l'objet d'une immunosurveillance fonctionnelle, et le privilège immunitaire peut être un critère clé pour identifier les CSCs. En outre, le modèle suggère que les CSCs peuvent initialement dépendre de niches de cellules souches, et les CSCs peuvent y fonctionner comme un réservoir dans lequel des mutations peuvent s'accumuler pendant des décennies sans être limitées par le système immunitaire. Des tumeurs peuvent spontanément se développer si: A) les CSCs perdent leur dépendance vis-à-vis des niches de CS (tumeurs moins différenciées), B) leur progéniture de cellules tumorales normales hautement prolifératives, mais initialement immunogènes, développe des moyens d'échapper à l'immunosurveillance ou, C) le système immunitaire peut perdre sa capacité de suppression des tumeurs, par exemple en raison du vieillissement.
Débat
L'existence des CSCs fait l'objet de débats, car de nombreuses études n'ont trouvé aucune cellule présentant leurs caractéristiques spécifiques. Les cellules cancéreuses doivent être capables de proliférer et de se renouveler en permanence afin de conserver les nombreuses mutations nécessaires à la cancérogenèse et soutenir la croissance d'une tumeur. En effet, les cellules différenciées (limitées par la limite de Hayflick) ne peuvent pas se diviser indéfiniment. Sur le plan thérapeutique, si la plupart des cellules tumorales sont dotées de propriétés de cellules souches, cibler directement la taille de la tumeur est une stratégie valable. Si les CSCs représentent une petite minorité, les cibler peut être plus efficace. Un autre débat porte sur l'origine des CSCs - qu'il s'agisse de la dérégulation des cellules souches normales ou d'une population plus spécialisée qui a acquis la capacité de se renouveler elle-même (ce qui est lié à la question de la plasticité des cellules souches). La découverte d’une plasticité phénotypique des cellules cancéreuses induite par la thérapie, altérant leurs transcriptomes et leur conférant des propriétés de type cellule souche pour échapper à la destruction, vient troubler ce débat.
Mise en évidence
Les premières preuves concluantes concernant l’existence des CSC sont apparues en 1997. Bonnet et Dick ont isolé une sous-population de cellules leucémiques qui exprimaient le marqueur de surface CD34, mais pas CD38. Les auteurs ont établi que la sous-population CD34+/CD38− est capable d'initier des tumeurs chez les souris NOD/SCID qui étaient histologiquement similaires au donneur. La première preuve de l'existence de cellule souche cancéreuse dans une tumeur solide a suivi en 2002 avec la découverte d'un clonogène, une cellule sphérique isolée et caractérisée à partir de gliomes de cerveau humain adulte. Les tumeurs gliales corticales humaines contiennent des cellules souches neurales exprimant des marqueurs astrogliaux et neuronaux in vitro. Il a été démontré que les cellules souches cancéreuses isolées de gliomes humains adultes induisent des tumeurs qui ressemblent à la tumeur mère lorsqu'elles sont greffées sur des modèles intracrâniens de souris nude.
Dans les expériences de recherche sur le cancer, des cellules tumorales sont parfois injectées à un animal de laboratoire pour établir une tumeur. La progression de la maladie est alors suivie dans le temps et l'efficacité des nouveaux médicaments peut être testée. La formation d'une tumeur nécessite l'introduction de milliers ou de dizaines de milliers de cellules. Classiquement, cela s'expliquait par une mauvaise méthodologie (c'est-à-dire que les cellules tumorales perdent leur viabilité lors du transfert) ou par l'importance critique du micro-environnement, l'environnement biochimique particulier des cellules injectées. Les partisans du paradigme des CSC affirment que seule une petite fraction des cellules injectées, les CSC, a le potentiel de générer une tumeur. Dans la leucémie aiguë myéloïde humaine, la fréquence de ces cellules est inférieure à 1 sur 10 000.
D'autres preuves proviennent de l'histologie. De nombreuses tumeurs sont hétérogènes et contiennent de multiples types de cellules natives de l'organe hôte. L'hétérogénéité de la tumeur est généralement conservée par les métastases de la tumeur. Cela suggère que la cellule qui les a produites avait la capacité de générer de multiples types de cellules, une caractéristique classique des cellules souches.
L'existence des cellules souches de la leucémie a incité la recherche sur d'autres cancers. Des CSC ont été identifiées dans plusieurs tumeurs solides, notamment:
- Cerveau
- Sein
- Côlon
- Ovaire
- Pancréas
- Prostate
- Mélanome
- Myélome Multiple
- Cancer de la peau non mélanome
Modèles mécanique et mathématique
Après avoir émis des hypothèses sur les mécanismes conduisant au cancer, il est possible de développer des modèles mathématiques prédictifs en se basant sur la méthode du compartiment cellulaire. Par exemple, les croissances de cellules anormales peuvent être désignées par des probabilités de mutations spécifiques. Un tel modèle prédit qu’une succession d’altérations génétiques des cellules matures augmente la formation d'une progéniture anormale et le risque de cancer. L'efficacité clinique de tels modèles reste encore à établir.
Origine

L'origine des CSCs est un domaine de recherche actif. La réponse peut dépendre du type de tumeur et du phénotype. Jusqu'à présent, l'hypothèse selon laquelle les tumeurs proviennent d'une seule "cellule d'origine" n'a pas été démontrée à l'aide du modèle des cellules souches cancéreuses. Cela s'explique par le fait que les cellules souches cancéreuses ne sont pas présentes dans les tumeurs en phase terminale.
Les hypothèses sur l’origine des CSCs comprennent des mutants dans des cellules souches progénitrices ou en développement, des mutants dans des cellules souches adultes ou des cellules progénitrices adultes et des mutants dans des cellules différenciées qui acquièrent des attributs de type souches. Ces théories se concentrent souvent sur la "cellule d'origine" d'une tumeur.
Hypothèses
Mutation des cellules souches
L'hypothèse de la "mutation des populations de niche de cellules souches au cours du développement" affirme que ces populations de cellules souches en développement subissent une mutation puis se multiplient de sorte que la mutation est partagée par de nombreux descendants. Ces cellules filles sont beaucoup plus proches de devenir des tumeurs et leur nombre augmente la probabilité d'une mutation cancéreuse.
Cellules souches adultes
Une autre théorie associe les cellules souches adultes (CSA) à la formation de tumeurs. Cette dernière est le plus souvent associée à des tissus présentant un taux élevé de renouvellement cellulaire (comme la peau ou l'intestin). Dans ces tissus, les CSA représentent des cibles idéales en raison de leurs fréquentes divisions cellulaires (par rapport à la plupart des CSA) et de leur longue durée de vie. Cette combinaison crée un ensemble de circonstances favorables à l'accumulation de mutations : l'accumulation de mutations est le principal facteur qui entraîne l'apparition du cancer. Les preuves montrent que cette association représente un phénomène réel, bien que des cancers spécifiques aient été liés à une cause précise.
Dé-différenciation
La dé-différenciation des cellules mutées peut créer des caractéristiques semblables à celles des cellules souches, ce qui suggère que n'importe quelle cellule peut devenir une cellule souche cancéreuse. En d'autres termes, une cellule entièrement différenciée subit des mutations ou des signaux extracellulaires qui la ramènent à un état de cellule souche. Ce concept a été démontré tout récemment dans des modèles de cancer de la prostate. Le traitement par privation androgénique semble altérer transitoirement le transcriptome des cellules en cellule souche de type crête neurale, avec les propriétés invasives et multipotentes de cette classe de cellules souches.
Hiérarchie
Le concept de hiérarchie des tumeurs affirme qu'une tumeur est une population hétérogène de cellules mutantes, qui partagent toutes certaines mutations, mais varient en fonction d'un phénotype spécifique. Une tumeur héberge plusieurs types de cellules souches, des cellules présentant un avantage sélectif pour l'environnement spécifique et d'autres cellules moins prospères. Ces lignées secondaires peuvent être plus avantagées dans d'autres environnements, ce qui permet à la tumeur de s'adapter, y compris à une intervention thérapeutique. Si ce concept est correct, il a une incidence sur les régimes de traitement spécifiques des cellules souches cancéreuses. Une telle hiérarchie compliquerait les tentatives visant à en déterminer l'origine.
Identification
Les CSCs, qui sont maintenant signalées dans la plupart des tumeurs humaines, sont couramment identifiées et enrichies à l'aide de stratégies d'identification des cellules souches normales qui sont similaires d'une étude à l'autre. Ces procédures comprennent le tri des cellules activées par fluorescence (FACS), avec des anticorps dirigés contre les marqueurs de surface cellulaire et des approches fonctionnelles comprenant un test permettant d’isoler une population cellulaire appelée "side population » ou un test Aldefluor. Les populations enrichies en CSCs sont ensuite implantées, à différentes doses, chez des souris immunodéficientes pour évaluer leur capacité de développement tumoral. Ce test in vivo est appelé test de dilution limite. Les sous-ensembles de cellules tumorales qui peuvent initier le développement de la tumeur à un faible nombre de cellules sont ensuite testés pour leur capacité d'auto-renouvellement dans des études tumorales en série.
Les CSCs peuvent également être identifiées par une coloration Hoechst. Cette technique est basée sur l’efflux du colorant via des transporteurs membranaires de multiples drogues (MDR) et de la famille ABC (ATP binding cassettes).
Une autre approche consiste à effectuer des tests de sphères. De nombreuses cellules souches normales, comme les cellules hématopoïétiques ou les cellules souches de tissus, dans des conditions de culture spéciales, forment des sphères tridimensionnelles qui peuvent se différencier. Comme les cellules souches normales, les CSCs isolées de tumeurs du cerveau ou de la prostate ont également la capacité de former des sphères indépendantes d'ancrage.
Hétérogénéité (marqueurs)
Des CSCs ont été identifiées dans diverses tumeurs solides. Généralement, des marqueurs spécifiques aux cellules souches normales sont utilisés pour isoler les CSCs des tumeurs solides et hématologiques. Les marqueurs les plus fréquemment utilisés pour l'isolement des CSCs sont les suivants : CD133 (également appelé PROM1), CD44, ALDH1A1, CD34, CD24 et EpCAM (molécule d'adhésion des cellules épithéliales, également appelée antigène spécifique de l'épithélium, ESA).
CD133 (prominine 1) est une glycoprotéine à cinq domaines transmembranaires exprimée par les cellules souches et progénitrices CD34+, dans les précurseurs endothéliaux et les cellules souches neurales fœtales. Elle a été détectée grâce à son épitope glycosylé appelé AC133.
EpCAM (molécule d'adhésion des cellules épithéliales , ESA, TROP1) est une molécule d'adhésion cellulaire hémophile indépendante de Ca2+, exprimée sur la surface basolatérale de la plupart des cellules épithéliales.
CD90 (THY1) est une glycosylphosphatidylinositol glycoprotéine ancrée dans la membrane plasmique et impliquée dans la transduction du signal. Elle peut également jouer un rôle de médiateur dans l'adhésion entre les thymocytes et le stroma thymique.
CD44 (PGP1) est une molécule d'adhésion qui joue un rôle pléiotropique dans la signalisation, la migration et le guidage des cellules. Elle a de multiples isoformes, dont CD44H, qui présente une grande affinité pour l'hyaluronate, et CD44V, qui a des propriétés métastatiques.
CD24 (HSA) est une molécule d'adhésion glycosylée fixée au glycosylphosphatidylinositol , qui joue un rôle de co-stimulation dans les cellules B et T.
CD200 (OX-2) est une glycoprotéine membranaire de type 1, qui délivre un signal inhibiteur aux cellules immunitaires, dont les cellules T, les cellules tueuses naturelles (NK, natural killer) et les macrophages.
Les ALDH sont une famille d'enzymes omniprésentes de la famille des aldéhydes déshydrogénases, qui catalysent l'oxydation des aldéhydes aromatiques en acides carboxyliques. Par exemple, elles jouent un rôle dans la conversion du rétinol en acide rétinoïque, qui est essentiel à la survie.
La première tumeur maligne solide à partir de laquelle les CSCs ont été isolées et identifiées était le cancer du sein. Les CSCs issues de cancer du sein sont les plus étudiées. Les CSCs du sein ont été enrichies en sous-populations CD44+CD24-/low, SP et ALDH+. Les CSCs du sein présentent donc des phénotypes divers. L'expression des marqueurs des CSCs dans les cellules de cancer du sein est apparemment hétérogène et les populations de CSCs du sein varient selon les tumeurs. Les populations de cellules CD44+CD24− et CD44+CD24+ sont toutes deux des cellules initiatrices de tumeurs ; cependant, les CSCs sont plus fortement enrichies en utilisant le profil de marqueur CD44+CD49fhiCD133/2hi.
Des CSCs ont été identifiées dans de nombreuses tumeurs cérébrales. Les cellules tumorales de type souches ont été identifiées à l'aide de marqueurs de surface cellulaire, notamment CD133, SSEA-1 (stage-specific embryonic antigen-1), EGFR et CD44. Dans les tumeurs cérébrales, l'utilisation de CD133 pour l'identification de cellules souches peut être problématique. D’une part, les cellules tumorigènes se trouvent à la fois dans les populations de cellules CD133+ et CD133− dans certains gliomes. D’autre part, certaines cellules CD133+ peuvent ne pas posséder de capacité d'initiation tumorale.
Des CSCs ont été décrites dans le cancer du côlon. Pour les identifier, des marqueurs de surface cellulaire tels que CD133, CD44 et ABCB5, l'analyse fonctionnelle, ainsi que l'analyse clonale et le test Aldefluor ont été utilisés. L'utilisation de CD133 comme marqueur positif pour les CSCs du côlon a donné des résultats contradictoires. L'épitope AC133, mais pas la protéine CD133, est spécifiquement exprimé dans les CSCs du côlon et son expression est perdue lors de la différenciation. En outre, les cellules CD44+ du cancer du côlon et une sous-fraction supplémentaire de la population de cellules CD44+EpCAM+ avec CD166 améliorent le succès des greffes de tumeurs.
De multiples CSCs ont été signalées dans la prostate, les poumons et de nombreux autres organes, notamment le foie, le pancréas, les reins ou les ovaires. Dans le cancer de la prostate, les cellules initiatrices de tumeur ont été identifiées dans le sous-ensemble de cellules CD44+ comme étant des cellules CD44+α2β1+, TRA-1-60+CD151+CD166+ ou ALDH+. Des marqueurs putatifs pour les CSCs pulmonaires ont été décrits, notamment CD133+, ALDH+, CD44+ et la protéine onco-fœtale 5T4+.
Métastases
Les métastases sont la cause principale de la létalité des tumeurs. Cependant, toutes les cellules tumorales ne peuvent pas métastaser. Ce potentiel dépend de facteurs qui déterminent la croissance, l'angiogenèse, l'invasion et d'autres processus fondamentaux.
Transition épithélio-mésenchymateuse
Dans les tumeurs épithéliales, la transition épithélio-mésenchymateuse (TEM) est considérée comme un événement crucial. La TEM et la transition inverse, à savoir la transition mésenchymateuse-épithéliale (TME), sont impliquées dans le développement embryonnaire, qui implique la perturbation de l'homéostasie des cellules épithéliales et l'acquisition d'un phénotype mésenchymateux migrateur. La TEM semble être contrôlée par des voies canoniques telles que la voie WNT et la voie du TGFb (le facteur de croissance transformant β).
Une caractéristique importante de la TEM est la perte de l’expression membranaire de la E-cadhérine au niveau des jonctions adhérentes, où la β-caténine pourrait jouer un rôle important. La translocation de la ß-caténine des jonctions adhérentes vers le noyau peut entraîner une perte de la E-cadhérine et, par conséquent, une TEM. La β-caténine nucléaire peut apparemment activer directement, par transcription, les gènes cibles associés à la TEM, tels que SLUG (également connu sous le nom de SNAI2), répresseur transcriptionnel du gène codant la E-cadhérine. Les propriétés mécaniques du microenvironnement de la tumeur, comme l'hypoxie, peuvent contribuer à la survie et au potentiel métastatique de la CSC par la stabilisation des facteurs induits par l'hypoxie grâce aux interactions avec les dérivés réactifs de l’oxygène (ROS, reactive oxygen species).
Les cellules tumorales soumises à une TEM peuvent être des précurseurs de cellules cancéreuses métastatiques, voire de CSCs métastatiques. Dans la bordure invasive du carcinome pancréatique, un sous-ensemble de cellules CD133+CXCR4+ (récepteur de la chimiokine CXCL12 également connu sous le nom de ligand SDF1) a été défini. Ces cellules présentaient une activité migratoire nettement plus forte que leurs homologues CD133+CXCR4−, mais toutes deux avaient une capacité de développement tumoral similaire. De plus, l'inhibition du récepteur CXCR4 a réduit le potentiel métastatique sans altérer la capacité tumorigène.
Motif d’expression en deux phases
Dans le cancer du sein, les cellules CD44+CD24−/low sont détectables dans les épanchements pleuraux métastatiques. En revanche, un nombre accru de cellules CD24+ a été identifié dans les métastases distantes chez les patientes atteintes d'un cancer du sein. Il est possible que les cellules CD44+CD24−/low se détachent de la tumeur initiale pour former des métastases et une fois dans leur nouveau site, elles changent de phénotype et subissent une différenciation limitée. L'hypothèse du schéma d'expression en deux phases propose deux formes de cellules souches cancéreuses : stationnaires (CSS) et mobiles (CSM). Les CSS sont ancrées dans les tissus et persistent dans les zones différenciées tout au long de la progression de la tumeur. Les CSM sont situées à l'interface tumeur-hôte. Ces cellules sont apparemment dérivées des CSS à travers l'acquisition d'une TEM transitoire (Fig. 7).

Applications
Le ciblage des CSCs jouent un rôle dans le traitement contre le cancer, notamment à travers l'identification de la maladie, la sélection de médicaments cibles, la prévention des métastases et les stratégies d'intervention.
Traitement
Les CSCs sont intrinsèquement plus résistantes aux agents chimiothérapeutiques. Cinq facteurs principaux y contribuent :
- 1. Leur niche les protège contre le contact avec de grandes concentrations de médicaments anticancéreux;
- 2. Elles expriment diverses protéines transmembranaires, telles que MDR1 et BCRP, qui pompent les médicaments hors du cytoplasme;
- 3. Elles se divisent lentement, comme les cellules souches adultes ont tendance à le faire, et ne sont donc pas tuées par les agents chimiothérapeutiques qui ciblent les cellules qui se répliquent rapidement en endommageant l'ADN ou en inhibant la mitose;
- 4. Elles régulent les protéines de réparation des dommages à l'ADN;
- 5. Elles sont caractérisées par une suractivation des voies de signalisation anti-apoptotiques.
Après une chimiothérapie, les CSCs ayant résisté au traitement sont capables de repeupler la tumeur et de provoquer une rechute. Un traitement supplémentaire visant à éliminer les CSCs en plus des cellules somatiques cancéreuses doit être utilisé pour empêcher cela.
Ciblage (Thérapie ciblée)
Le ciblage sélectif des CSCs peut permettre de traiter des tumeurs agressives et non résécables, ainsi que de prévenir les métastases et les rechutes. L'hypothèse suggère que lors de l'élimination des CSCs, le cancer pourrait régresser en raison de la différenciation et/ou de la mort cellulaire. La fraction des cellules tumorales qui sont des CSCs et qui doivent donc être éliminées reste floue.
Des études ont cherché à identifier des marqueurs spécifiques et des signatures tumorales protéomiques et génomiques qui distinguent les CSCs des autres cellules. En 2009, une équipe de scientifiques a montré que la salinomycine pouvait réduire de façon sélective et par plus de 100 fois la proportion de CSCs du sein chez des souris par rapport au Paclitaxel, un agent chimiothérapeutique couramment utilisé. Certains types de cellules cancéreuses peuvent survivre au traitement à la salinomycine par autophagie. C'est-à-dire que les cellules utilisent des organites acides tels que les lysosomes pour dégrader et recycler certains types de protéines. L'utilisation d'inhibiteurs de l’autophagie peut tuer les cellules souches cancéreuses qui survivent grâce à ce processus.
Le récepteur de surface cellulaire interleukine-3-alpha (CD123) est surexprimé sur les cellules souches leucémiques (CSL) CD34+CD38− dans la leucémie aiguë myéloblastique (LAM) mais pas sur les cellules normales CD34+CD38− de la moelle osseuse. Le traitement de souris NOD/SCID ayant subi une greffe de LAM, par un anticorps monoclonal anti-CD123 altère le foyer de CSLs au niveau de la moelle osseuse et réduit la prolifération globale des cellules de LAM ainsi que la proportion de CSLs chez les souris receveuses.
Une étude réalisée en 2015 a permis de conditionner des nanoparticules avec du miR-34a et du bicarbonate d'ammonium et de les administrer à des CSCs de prostate dans un modèle de souris. Ensuite, les chercheurs ont irradié la zone avec une lumière laser proche de l’infrarouge. Ceci conduit à un gonflement d’au moins trois fois plus la taille des nanoparticules, faisant éclater les endosomes et dispersant l'ARN dans la cellule. Le miR-34a peut faire baisser les niveaux de CD44.
Une étude de 2018 a identifié des inhibiteurs de la famille des enzymes ALDH1A et a montré qu'ils pouvaient dépléter sélectivement les cellules souches cancéreuses putatives dans plusieurs lignées cellulaires de cancer de l'ovaire.
Voies de signalisation
Le développement de nouveaux médicaments ciblant les CSCs nécessite une meilleure compréhension des mécanismes cellulaires qui régulent la prolifération cellulaire. Les premiers progrès dans ce domaine ont été réalisés avec les cellules souches hématopoïétiques (CSH) et leurs homologues transformés dans la leucémie, la maladie pour laquelle l'origine des CSCs est la mieux comprise. Les cellules souches de nombreux organes partagent les mêmes voies cellulaires que les CSHs issues de leucémies.
Une cellule souche normale peut être transformée en CSC en dérégulant les voies de prolifération et de différenciation qui la contrôlent ou en induisant une activité oncoprotéique.
BMI-1
Le répresseur transcriptionnel du groupe Polycomb, Bmi-1, a été découvert comme un oncogène commun activé dans les lymphomes et a ensuite été montré comme régulant les CSHs. Le rôle de Bmi-1 a été illustré dans les cellules souches neurales. La voie semble être active dans les CSCs des tumeurs cérébrales pédiatriques.
Notch
La voie Notch joue un rôle dans le contrôle de la prolifération des cellules souches pour plusieurs types de cellules, notamment les CS hématopoïétiques, neurales et mammaires. Des composants de cette voie ont été proposés pour agir comme oncogènes dans les tumeurs mammaires et autres.
Une branche de la voie de signalisation Notch qui implique le facteur de transcription Hes3 régule en culture le nombre de cellules de glioblastome qui présentent les caractéristiques de CSCs.
Sonic hedgehog et Wnt
Ces voies de développement sont des régulateurs des cellules souches (CS). Les voies Sonic hedgehog (SHH) et Wnt sont toutes deux couramment hyperactivées dans les tumeurs et sont nécessaires pour soutenir la croissance tumorale. Toutefois, les facteurs de transcription Gli qui sont régulés par SHH tirent leur nom des gliomes, où ils sont fortement exprimés. Il existe un certain degré d’interférence entre les deux voies et elles sont généralement activées ensemble. En revanche, dans le cancer du côlon, la voie de signalisation Hedgehog semble antagoniser la voie Wnt.
Il existe des inhibiteurs de la voie Sonic hedgehog, comme la cyclopamine. La cyclopamine sous sa forme soluble peut être plus efficace dans le traitement du cancer. Le diméthylaminoparthénolide (DMAPT), un dérivé hydrosoluble du parthénolide, induit un stress oxydatif et inhibe la signalisation NF-κB dans la LAM (leucémie) et éventuellement dans le myélome et le cancer de la prostate. La télomérase est un sujet d'étude dans la physiologie des CSCs. Récemment, un essai clinique évaluant l’effet du GRN163L (Imetelstat) sur le ciblage des cellules souches du myélome a débuté.
La signalisation Wnt peut devenir indépendante de stimuli réguliers, à travers des mutations dans les oncogènes en aval et les gènes suppresseurs de tumeurs qui sont activés de façon permanente même si le récepteur normal n'a pas reçu de signal. La β-caténine se lie à des facteurs de transcription tels que la protéine TCF4 et, en combinaison, les molécules activent les gènes nécessaires. Le LF3 inhibe fortement la liaison de la ß-caténine in vitro, dans les lignées cellulaires et réduit la croissance des tumeurs dans des modèles de souris. Il empêche la réplication et réduit leur capacité à migrer, le tout sans affecter les cellules saines. Après le traitement par le LF3, il ne restait plus de cellules souches cancéreuses. Cette découverte est le fruit d'une "conception rationnelle des médicaments", faisant appel aux technologies AlphaScreens et ELISA.
La voie Wnt/ß-caténine est régulièrement activée dans les cellules cancéreuses conduisant à l’expression de gènes cible de façon constitutive. L’un des gènes activés, GAST, code hPG80 ou progastrine circulante dans le sang. Initialement considérée comme biologiquement inactive, la hPG80 s'est avérée participer à certaines caractéristiques d'une tumeur, telles que la perturbation des jonctions cellule-cellule, la prolifération cellulaire, l'inhibition de l'apoptose, la régulation des cellules souches cancéreuses et l'angiogenèse. En outre, il a été démontré que le ciblage de la hPG80 par un anticorps spécifique favorise l'apoptose, diminue la prolifération et la migration des cellules cancéreuses colorectales humaines, et inhibe l'auto-renouvellement des CSCs de différentes origines, ainsi que la tumorigenèse Wnt chez la souris. Plus récemment il a été démontré que la hPG80 était exprimée par différents types de cancers même à des stades précoces.
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Cancer Stem Cell » (voir la liste des auteurs).
Références
Lectures complémentaires
- Rajasekhar Vinagolu K., Cancer Stem Cells, Hoboken, New Jersey, Wiley, , 552 p. (ISBN 978-1-118-35616-6)
- Cyril Corbet et Alexandre Prieur, Therapeutic Targeting of Cancer Stem-Like Cells (CSC) – The Current State of the Art, Frontiers Media SA, (ISBN 978-2-88963-647-1, lire en ligne)