

La barrière hémato-encéphalique, ou hémo-encéphalique, ou hémato-méningée est une barrière physiologique présente dans le cerveau chez tous les tétrapodes (vertébrés terrestres), entre la circulation sanguine et le système nerveux central (SNC). Elle sert à réguler le milieu (homéostasie) dans le cerveau, en le séparant du sang. Les cellules endothéliales, qui sont reliées par des jonctions serrées et qui tapissent les capillaires du côté du flux sanguin, sont les composants essentiels de cette barrière.
La barrière hémato-encéphalique protège le cerveau des agents pathogènes, des toxines et des hormones circulant dans le sang. Elle représente un filtre extrêmement sélectif, à travers lequel les nutriments nécessaires au cerveau sont transmis, et les déchets sont éliminés. Ce processus d'alimentation et d'élimination est produit par toute une série de mécanismes de transport actif.
Cette fonction de protection du cerveau complique le traitement médicamenteux d'un grand nombre de maladies neurologiques, car de nombreuses molécules actives ne peuvent pas traverser la barrière hémato-encéphalique. La recherche sur la manière de surmonter la barrière hémato-encéphalique est tout à fait actuelle. Bien peu de maladies – rares en plus – sont spécifiques de la barrière hémato-encéphalique, tandis qu'elle peut être atteinte par de nombreuses maladies générales. Une atteinte, ou une lésion, de la barrière hémato-encéphalique est une complication à prendre très au sérieux.
Les premières expériences, qui ont indiqué l'existence de cette barrière ont été conduites par Paul Ehrlich en 1885. Mais il a mal interprété les résultats de ses expériences. La preuve définitive de l'existence de la barrière n'a été donnée qu'en 1967 par des recherches en microscopie électronique en transmission.
Fonctions de la barrière hémato-encéphalique



Chez l'humain, le cerveau représente environ 2 % de la masse corporelle. Mais ses besoins en énergie sont environ de 20 % du total. Contrairement aux autres organes du corps, le cerveau dispose de très peu de réserves en nutriments et en oxygène. Et les cellules nerveuses ne sont pas capables de satisfaire leurs besoins en énergie de manière anaérobie, c'est-à-dire sans aucun apport d'oxygène élémentaire. C'est ainsi qu'une interruption de l’apport de sang au cerveau amène au bout de 10 s une syncope (perte de connaissance), et quelques minutes après, les cellules nerveuses commencent à mourir. Selon l’activité de chaque domaine du cerveau, ses besoins en énergie et ses réserves peuvent être très différents. Pour ajuster les apports aux besoins, chaque domaine est en mesure de régler par lui-même les apports sanguins qui lui sont nécessaires.
Les fonctions complexes du cerveau sont liées à des processus électrochimiques et biochimiques très sensibles, qui ne peuvent se dérouler que dans un milieu interne homéostatique largement débarrassé de toutes perturbations. Par exemple, les oscillations du pH du sang (une mesure du caractère basique ou acide) ne doivent pas se répercuter sur le cerveau. Les variations de la concentration en potassium changeraient le potentiel de la membrane des cellules nerveuses. Les neurotransmetteurs emportés par le sang dans les vaisseaux ne doivent pas pénétrer dans le système nerveux central, car ils y perturberaient sérieusement le fonctionnement des synapses qui s'y trouvent. En plus, les neurones ne sont pas capables de se régénérer en cas de dommage dû à une variation du milieu. Enfin, le cerveau, organe de commande central, doit être protégé de l’influence de matières étrangères au corps, telles que par exemple des xénobiotiques, ou des agents pathogènes. L'imperméabilité considérable de la barrière hémato-encéphalique à l'égard des agents pathogènes, des anticorps et des leucocytes en fait une « barrière immunologique ».
Par ailleurs, en raison des besoins très importants en énergie du cerveau – par comparaison avec d'autres organes – des quantités de déchets biochimiques très importantes doivent être éliminées à travers la barrière hémato-encéphalique.
Pour accomplir toutes ces fonctions (alimentation, élimination et homéostasie), le circuit des vaisseaux sanguins cérébraux des vertébrés présente, par comparaison avec les vaisseaux périphériques, toute une série de différences structurelles et fonctionnelles. Cette différenciation exerce une très large séparation du cerveau de l'espace extracellulaire environnant, et est une condition essentielle pour la protection du tissu neuronal sensible, et pour l'obtention d'un milieu interne stable.
Les changements du fonctionnement de la barrière hémato-encéphalique provoquent des altérations du système nerveux central, et peuvent en provoquer des troubles fonctionnels ou des maladies. Par suite, une série de maladies neurologiques est reliée plus ou moins directement à la barrière hémato-encéphalique.
Anatomie de la barrière hémato-encéphalique

L'élément essentiel de la barrière hémato-encéphalique est constitué par les cellules endothéliales avec leurs jonctions serrées. Mais deux autres types de cellules sont également importants, tant du point de vue de la fonction que celui de la naissance et de la croissance de la barrière hémato-encéphalique : les péricytes et les astrocytes. Les interactions entre cellules endothéliales, péricytes et astrocytes sont plus étroites qu'entre tous autres types de cellules. Ces trois types de cellules forment ensemble la barrière hémato-encéphalique de la plupart des vertébrés, la barrière hémato-encéphalique cellulaire. Il existe dans le règne animal d'autres types de barrière hémato-encéphalique, qui sont abordés dans l'article détaillé.
Endothélium
Les capillaires sont tapissés – comme les vaisseaux périphériques – de cellules endothéliales. Dans le cerveau, celles-ci ont une structure particulièrement étanche. le nombre des mitochondries est environ de 5 à 10 fois plus élevé que dans les capillaires périphériques, en raison de l'énergie nécessitée pour exercer un transport actif des nutriments nécessaires à travers les cellules. Les cellules endothéliales présentent sur leurs membranes une quantité d'aquaporines, canaux spécialisés pour le passage de l’eau, pour la régulation de la quantité d'eau au sein du cerveau.
L'étanchéité de la barrière peut être quantifiée par sa résistance électrique. Chez un rat adulte, la résistance monte à environ 2 000 Ω cm2. Dans les capillaires musculaires, elle n'est que d'environ 30 Ω cm2.
Jonctions serrées

Les cellules endothéliales sont liées ensemble par de solides liaisons, appelées jonctions serrées, qui rendent imperméable l'espace entre cellules. Plusieurs types de protéines membranaires les ceinturent afin d'assurer l'étanchéité.
Lame basale
Les cellules endothéliales sont entourées par une couche protéique, la lame basale épaisse de 40 à 50 nm, donc visible seulement au microscope électronique.
Péricytes
Les péricytes sont de petites cellules ovales, qui couvrent en tout 20 % de la surface externe des capillaires, solidement amarrées aux cellules endothéliales. Elles jouent trois rôles principaux :
- Un rôle moteur permis par leur haut contenu en actine, et qui module la section du capillaire en fonction des besoins.
- Un rôle de macrophage, qui leur permet d'intervenir en seconde ligne de défense contre les agressions venant de la circulation sanguine.
- Un rôle de régulateur des divisions cellulaires et de la différenciation cellulaire des cellules endothéliales. Ils jouent notamment un rôle important lors de la formation de nouveaux vaisseaux (angiogenèse).
Astrocytes

Les astrocytes sont des cellules en étoile, significativement plus grandes que les péricytes. Ils couvrent les capillaires du cerveau à 99 % avec leurs pieds enchevêtrés en rosettes. L'interaction immédiate (20 nm) entre cellules endothéliales et astrocytes induit dans les deux sens les spécificités anatomiques.
Leurs fonctions principales sont :
- une modulation rapide de la perméabilité des cellules endothéliales,
- l'alimentation des neurones,
- la régulation du milieu extracellulaire,
- la synthèse du cholestérol, qui ne peut pas traverser la barrière hémato-encéphalique, pour la myéline des gaines des axones neuronaux
Domaines du cerveau sans barrière hémato-encéphalique
Tous les capillaires du cerveau ne font pas partie de la barrière hémato-encéphalique : les parties du cerveau qui sécrètent des hormones et celles qui ont une fonction sensorielle sur la composition du sang doivent rester en communication avec la circulation sanguine.
On dénombre six organes circumventriculaires partiellement démunis de barrière hémato-encéphalique. Il s'agit de l'organe subfornical (en), l'organe vasculaire de la lame terminale, la neurohypophyse, la glande pinéale (ou épiphyse), l'organe subcommissural et l'area postrema. Ces régions sont entourées de tanycytes, analogues aux épendymocytes qui séparent le cerveau du liquide cérébrospinal remplissant l'épendyme, mais avec des jonctions serrées très étanches.
Autres informations
On consultera avec profit l'article détaillé pour des informations sur :
- Les données générales et statistiques sur la barrière hémato-encéphalique
- Les phases du développement de la barrière chez le fœtus et le nouveau-né
- Des perspectives sur l'évolution de la barrière chez les vertébrés supérieurs, et les paradoxes qu'elle présente.
Barrière sang - liquide cérébrospinal
Outre la barrière hémato-encéphalique, il existe une deuxième barrière entre la circulation sanguine et le système nerveux central : la barrière sang-LCS. Cette barrière est formée par les cellules épithéliales et les jonctions serrées des plexus choroïdes. La barrière sang-LCS a aussi une part de l'homéostasie du cerveau. Elle l'approvisionne en vitamines, en nucléotides et en glucose. La contribution au transport de matières vers le cerveau est en fin de compte assez faible, et totalement insuffisante pour couvrir les besoins du cerveau en nutriments et oxygène. La surface d'échange que forment les capillaires intracérébraux de la barrière hémato-encéphalique représente 5 000 fois celle des plexus choroïdes.
Outre ces deux barrières, si importantes pour le système nerveux central, on trouve dans le corps d'autres barrières ultrasélectives analogues, qui contrôlent l'échange de matières avec le sang. Parmi d'autres, ce sont :
- la barrière système nerveux central-liquide cérébrospinal, assurée principalement par les épendymocytes et autres cellules des plexus choroïdes. Les astrocytes assurent la communication entre les deux barrières par différents types de pieds ;
- la barrière sang-placenta ;
- la barrière entre le sang et les tubes séminifères, assurée par des jonctions serrées entre cellules de Sertoli ;
- la barrière entre sang et urine, assurée par à la fois par une limitation en taille des molécules pouvant traverser, et par une charge électrique négative des membranes, repoussant les protéines du sang ;
- la barrière entre sang et thymus, destinée à protéger les lymphocytes-T de tout contact avec des antigènes pendant leur maturation. Elle est effectuée par une succession de cinq couches différentes de cellules dans la paroi des capillaires
- la barrière des poumons : sang et air ne sont séparés que par deux couches de cellules, l'endothélium des capillaires et l'épithélium des poumons, partageant la même lame basale.
Processus de transport à la barrière hémato-encéphalique
La barrière hémato-encéphalique doit assurer, malgré son étanchéité, le transport de nutriments et d'oxygène vers le cerveau, et éliminer les déchets.
- Transport paracellulaire
Pour éviter toute fuite incontrôlée, les cellules endothéliales sont liées par des jonctions serrées étanches. Seules de toutes petites molécules peuvent passer à travers les jonctions serrées : eau, glycérine ou urée.
- Diffusion libre

La forme la plus simple est la diffusion libre ou passive, qui tend à établir un équilibre de concentration ou de potentiel chimique des substances. Elle ne requiert aucune énergie. Le débit est proportionnel à la différence de potentiel et n'est pas contrôlable.
Les petites molécules peuvent franchir la membrane par des trous correspondant à des déformations locales des chaînes de phospholipides constituant la membrane. Les trous sont mobiles, et peuvent donc accompagner la molécule dans son trajet à travers la membrane. Encore faut-il que la molécule en question ait une affinité raisonnable pour les lipides. Ce processus ne concerne donc essentiellement que les petites molécules lipophiles (hydrophobes).
- Passage par canaux

Les petites molécules polaires, comme l'eau, ne peuvent pratiquement pas diffuser à travers les membranes par le processus décrit. On trouve dans la membrane cellulaire un grand nombre de protéines qui jouent le rôle de canaux spécialisés pour le passage de l'eau : les aquaporines. Elles offrent une grande perméabilité à l'eau, dans les deux sens selon la différence de pression osmotique. Il existe de nombreux autres types de canaux, plus ou moins spécialisés, qui peuvent être ouverts ou fermés sous l'influence d'agents physiques. Mais tous ces canaux partagent la propriété de passivité : quand ils sont ouverts, ils laissent passer les molécules appropriées dans le sens de l'équilibre des concentrations.
- Diffusion facilitée
Des molécules vitales comme le glucose et certains acides aminés ne peuvent pas passer par des canaux. Il existe alors des transporteurs membranaires adaptés aux diverses molécules nécessaires. Les protéines membranaires de transport peuvent fonctionner comme uniport (une molécule à la fois), comme symport (deux molécules ou plus dans le même sens) ou comme antiport (deux molécules ou plus en sens contraires).
- Transport actif
Les transports décrits ci-dessus ne nécessitent de la part de la cellule aucune contribution énergétique. Mais il existe des substances qui doivent être transportées contre le gradient de concentration. Ceci nécessite alors une consommation d'énergie pour actionner des systèmes de transport actif ou « pompes ». Le transport du sang vers le cerveau est nommé « influx », et en sens inverse « efflux ». Certains de ces mécanismes sont très spécifiques, et identifient les molécules par leur forme, et distinguent donc les formes énantiomères gauche et droite. Par exemple, la D-asparagine est un ingrédient nécessaire pour la formation de certaines hormones. Elle bénéficie donc d'un transporteur actif d'influx. Par contre, la L-asparagine est un acide aminé stimulant dont l’accumulation dans le cerveau serait délétère. Elle est donc éliminée par un transport actif d'efflux.
Les transporteurs actifs d'efflux sont souvent peu spécifiques, leur rôle étant d'éliminer des déchets de nature parfois imprévisible.
Tous les types de transport pour tous les substrats n'ont pas encore été clairement identifiés.
- Transport vésiculaire

Les grosses molécules, ou même agrégats, qui ne peuvent pas utiliser de protéine membranaire de transport sont incorporées dans la cellule endothéliale par endocytose : la membrane plasmique se déforme en puits autour de l'objet à incorporer, puis la margelle du puits se soude, et la membrane recouvre son intégrité, tandis que l'objet est enfermé dans une vésicule. La vésicule peut traverser la cellule et s'ouvrir sur la face opposée par un mécanisme inverse, et libérer son contenu, c'est la transcytose.
- Transcytose à récepteurs
- S'il y a au puits de la membrane, des récepteurs qui se lient spécifiquement à la molécule visée, la vésicule est marquée, transportée et vidée. C'est le cas pour de grosses molécules comme la Lipoprotéine de basse densité (LDL), ingrédient de fabrication du cholestérol, l'insuline, et d'autres hormones peptidiques.
- Transcytose par adsorption
- Dans ce cas, la sélection se fait par la charge : le puits absorbe les molécules positivement chargées (les cations), d'où l'autre dénomination de « transport cationique ». Elle permet un plus grand débit que la transcytose à récepteurs.
- Principaux transporteurs
On consultera à ce sujet la table des principaux transporteurs.
Mesure et représentation de la perméabilité de la barrière hémato-encéphalique
Comme il est indiqué dans la section précédente, les processus de transport de substrats à travers la barrière hémato-encéphalique sont très variés, tant dans la nature du ou des substrats à transporter que dans le sens même où s'effectue le transport. Or il est essentiel pour la médecine et la pharmacie de savoir comment faire pénétrer dans le cerveau des médicaments (psychotropes) ou comment empêcher des toxiques, par exemple destinés à d'autres organes, d'y pénétrer.
La manière la plus classique est de procéder à des essais in vivo, sur l’animal, puis sur l'homme (« essais cliniques »), mais on peut recourir de façon plus aisée à des essais in vitro, voire à des simulations in silico.
Bases physiques
Un modèle simplifié, basé sur un seul capillaire a été mis au point par Renkin (1959) et Crone (1965). Le résultat s'exprime comme le « produit perméabilité-surface PS » de l’échantillon de capillaire. Il détermine la fraction E extraite en un passage d'une quantité de sang Q :
- .

Pour E < 0,2, la perméabilité est le facteur limitant, autrement elle est modérée ou grande.
Procédés in vitro
Le procédé le plus simple et le plus réaliste est l'utilisation de vaisseaux isolés, qui restent vivants pour un certain temps.
Avec des lignées de cellules endothéliales immortalisées, cultivées en couches simples, on peut faire des essais quantitatifs. La qualité de ces couches, celle des jonctions serrées, se mesure par leur résistance électrique, qui doit être aussi élevée que possible. Dans l'organisme vivant, elle peut être de l’ordre de 2 000 Ω cm2. Dans une culture mixte d'astrocytes et de cellules épithéliales, elle peut monter à 800 Ω cm2.
Procédés in vivo

Le premier procédé a été l'injection de colorants suivie de l'examen anatomique de l'animal. Le colorant franchissant la barrière hémato-encéphalique laisse une trace tenace. Ceci permet d'étudier des lésions volontaires de la barrière.
Les procédés in vivo sont irremplaçables pour leur sensibilité aux conditions physiologiques, le temps pendant lequel on peut les laisser agir et le nombre de passages du sang à travers le réseau capillaire.
- Indice d'absorption cérébrale
Le rapport entre les taux d'absorption d'une substance à tester et une substance facilement absorbée, toutes deux marquées radioactivement donne l'indice d'absorption cérébrale (Brain Uptake Index ou BUI). Cette méthode ne s'applique qu'à des substances rapidement absorbées. Voir la table pour quelques substances courantes.
- Indice d'efflux cérébral
Il est également intéressant de connaître pour chaque substrat les propriétés d'efflux de la barrière hémato-encéphalique. On compare le substrat testé à une matière de référence, peu capable de sortir de la barrière, toutes deux marquées radioactivement. Elles sont microinjectées directement dans le cerveau. L'indice d'efflux cérébral (Brain Efflux Index ou BEI) se calcule en fonction de ce qui reste de chacune des matières par rapport à ce qui a été injecté.
- Perfusion cérébrale
Dans la méthode de perfusion, le substrat marqué est longuement perfusé dans la carotide. Puis l'animal est sacrifié et la radioactivité du cerveau mesurée. Délicate, elle est réservée à des cas de BEI très faibles.
Il est intéressant de séparer les capillaires par centrifugation avant la mesure, afin d'éliminer tout le substrat qui lui est encore lié.
- Technique de diffusion d'indicateur
Dans cette technique, la substance de référence doit être incapable de traverser la barrière hémato-encéphalique. Le substrat à tester et la référence ne sont pas marqués radioactivement. Ils sont infusés dans la carotide, et dosés dans le sang de retour (veine jugulaire interne). Le dosage des matières permet le calcul de la quantité de substrat absorbée. Cette technique par différence ne convient donc que pour des substrats franchissant facilement la barrière.
- Autoradiographie quantitative
Voir dans le Wikilivre sur la photographie, les articles spécialisés sur l'autoradiographie et la fluorographie.

La figure ci-contre présente une autoradiographie d'un cerveau d'embryon de rat. Les domaines radioactifs sont foncés (zone subventriculaire SVZ). Le trait noir donne l'échelle de 2 mm.
Cette technique consiste en injection intraveineuse de substance marquée au carbone 14. Les organes sont disséqués, tranchés au microtome et déposés sur du film à rayons X. Connaissant la quantité de marqueur, on peut en déduire le produit perméabilité-surface de l’échantillon.
- Microdialyse intracérébrale
On implante dans le tissu nerveux une membrane hémiperméable. Par un microcathéter, on perfuse des substances, et/ou recueille le liquide interstitiel, éventuellement en continu.
En médecine humaine, la microdialyse intracérébrale est utilisée pour le monitoring neurochimique en cas d'accident vasculaire cérébral.
- Procédés d'imagerie
L'activité de la barrière hémato-encéphalique, le débit des capillaires, sont liés à l'activité du tissu nerveux qu'ils alimentent. On a donc une interaction entre ces trois grandeurs, qui peuvent varier substantiellement à l’échelle globale du cerveau. Ceci mène à dresser de façon non invasive des images globales du cerveau, essentiellement par trois méthodes qui se complètent : la tomographie par émission de positons (TEP), l'imagerie par résonance magnétique (IRM) et la spectroscopie par résonance magnétique (SRM).
- La tomographie par émission de positons
- La méthode repose sur des molécules marquées par un émetteur bêta+ : carbone 11 ou fluor 18. Le positon émis s'arrête dans la matière dense, et s'annihile avec un électron, donnant une paire de rayons gamma opposés. Le point de désintégration se situe donc sur la ligne joignant les points de détection des gammas. On peut ainsi, avec assez de désintégrations, reconstituer par le calcul la densité des molécules marquées.
- En raison de la courte demi-vie des émetteurs bêta+, cette méthode ne peut être utilisée qu'auprès de centres dotés de cyclotrons capables de fabriquer ces nucléides, et de laboratoires en mesure de les incorporer aux molécules à marquer.
- Imagerie par résonance magnétique (IRM)

- L'imagerie par résonance magnétique est trop peu sensible pour représenter le passage de substances actives à travers une barrière hémato-encéphalique saine. En cas de lésion, l’IRM avec produit de contraste joue un grand rôle.
- Spectroscopie par résonance magnétique (SRM)
- La SRM est une version de l'IRM où l'on fait continûment varier la fréquence pour exciter successivement divers noyaux, et donc leur réponse, ce qui se manifeste par un spectre avec des pics caractéristiques : fluor-19, carbone-13, phosphore-31, et hydrogène dans d'autres substances que l'eau. Les signaux très faibles nécessitent de longs temps de mesure et la mesure sur des volumes appréciables.
Stratégies pour franchir la barrière hémato-encéphalique
Comme il a été indiqué dans la section Processus de transport à la barrière hémato-encéphalique, il n'y a que peu de substances capables de franchir la barrière hémato-encéphalique, ce pourquoi beaucoup de médicaments psychotropes finissent par échouer à la barrière. 98 % de ces substances ne peuvent pas traverser la barrière hémato-encéphalique.
Il y a donc des dizaines d'années que l'on travaille intensément sur des méthodes susceptibles de rendre possible un transport de substance active dans le cerveau, en contournant – ou mieux en franchissant sélectivement – la barrière hémato-encéphalique. Un ensemble de stratégies pour surmonter la barrière hémato-encéphalique ont été mises au point dans ce but, ou en sont encore au stade d'élaboration.
En octobre 2014, la start-up française CarThera développe un dispositif novateur pour ouvrir temporairement la barrière hémato-encéphalique. Ce dispositif est basé sur utilisation conjointe de microbulles de gaz injectées dans la circulation sanguine et d'ultrasons focalisés. Le principe est le suivant : quand les ondes ultrasonores rencontrent des microbulles de gaz dans les vaisseaux sanguins aux abords du tissu biologique cible, celles-ci se mettent à osciller, entrainant alors des effets physiques et biologiques conduisant à la déstabilisation transitoire des cellules endothéliales de la barrière hématoencéphalique.
Dysfonctionnements de la barrière hémato-encéphalique
Des dysfonctionnements de la barrière hémato-encéphalique sont provoqués par le vieillissement, des blessures au cerveau, un stress et toutes sortes de pathologies. La barrière elle-même peut d'ailleurs être à l'origine de quelques maladies neurologiques très rares, de nature génétique.
Les perturbations du rôle protecteur de la barrière hémato-encéphalique sont une complication de beaucoup de maladies neurodégénératives et de blessures du cerveau. Certaines maladies périphériques, comme le diabète, ou certaines inflammations, ont une action nuisible sur le fonctionnement de la barrière hémato-encéphalique. La perte d'étanchéité de la barrière hémato-encéphalique a également été démontrée chez les patients souffrant d'épilepsie ou de la maladie d'Alzheimer.
D'autres pathologies peuvent perturber le fonctionnement des endothéliums « du dedans vers le dehors », c'est-à-dire que des influences provenant de la matrice extracellulaire perturbent l'intégrité de la barrière hémato-encéphalique. Par exemple, on a le glioblastome.
Mais un ensemble de maladies se manifestent dans le cerveau par le fait que certains agents peuvent pénétrer dans la barrière hémato-encéphalique. Parmi ceux-ci par exemple, le VIH, le virus T-lymphotrope humain, le virus du Nil occidental, certaines bactéries comme le méningocoque ou le vibrion cholérique.
Dans le cas de la sclérose en plaques, les agents pathogènes sont des cellules du système immunitaire de l'individu lui-même, qui franchissent la barrière hémato-encéphalique. De même, dans certains cancers non cérébraux, certaines cellules en métastase peuvent franchir la barrière hémato-encéphalique et donner lieu à des métastases cérébrales.
Un stress imposé à des souris (des épisodes de nage en eau froide) permet à des colorants (injectés en intraveineuse) de pénétrer dans le cerveau. De la même façon, la pénétration de la pyridostigmine dans le tissu cérébral participe au syndrome de la guerre du Golfe.
Agressions exogènes de la barrière hémato-encéphalique
Alcool
La consommation excessive d'alcool est un facteur majeur de risque pour les maladies psychophysiologiques, les inflammations et la sensibilité aux infections bactériennes. De plus, la consommation chronique d'alcool endommage la barrière hémato-encéphalique, ce qui est considéré comme un facteur important pour l'amorce de maladies neurodégénératives. Les dommages à la barrière hémato-encéphalique sont démontrés aussi bien par les recherches neuropathologiques sur les alcooliques que par des expériences sur des animaux.
Dans les expériences sur animaux, il a été établi que l'enzyme Myosin light-chain kinase (en) (MLCK) conduit dans l'endothélium à la phosphorylation de nombreuses protéines des jonctions serrées ou du cytosquelette des protéines, ce qui endommage l’intégrité de la barrière hémato-encéphalique. En outre, le stress oxydant dû à l'alcool conduit à des dommages supplémentaires à la barrière hémato-encéphalique.
Ce n'est pas l’alcool lui-même qui active l'enzyme MLCK dans l'endothélium, mais ses métabolites.
La dégradation fonctionnelle de la barrière hémato-encéphalique facilite la migration des leucocytes dans le cerveau, ce qui facilite le développement de pathologies neuro-inflammatoires.
Nicotine
L'abus chronique de nicotine sous forme de tabac fait non seulement augmenter le risque de cancer des poumons, mais aussi celui de maladie cardiovasculaire. Parmi les risques cardiovasculaires, il existe une corrélation directe avec les risques de démence. Plusieurs méta-analyses établissent que les fumeurs ont un risque significativement plus élevé de démence par maladie d'Alzheimer que les non-fumeurs. Le risque de démence vasculaire, ou de déficit cognitif léger n'est pas ou que peu augmenté. L'exposition quotidienne à la nicotine modifie chez les animaux non seulement la fonction mais aussi la structure de la barrière hémato-encéphalique des sujets. La substance modèle saccharose peut passer à travers les endothéliums significativement plus facilement, ce qui traduit en réalité une distribution modifiée des protéines de jonction serrée ZO-1, et une activité réduite de la claudine-3.
Après une exposition chronique à la nicotine, on a constaté dans l'endothélium une formation augmentée de microvillosités, de symport Na+/K+/2Cl− et pompe sodium-potassium déréglés.
Des études épidémiologiques, montrent que les fumeurs courent un risque significativement plus élevé de méningite bactérienne, par rapport aux non-fumeurs. La nicotine change les filaments d'actine du cytosquelette, ce qui semble faciliter le passage d'agents pathogènes comme le colibacille vers le cerveau.
Pour certains composés à diffusion limitée, par exemple l'antagoniste de la nicotine méthyllycaconitine qui se fixe sur le récepteur nicotinique à l'acétylcholine (nACHrs) et auquel on attribue des vertus pour le sevrage nicotinique, le passage de la barrière hémato-encéphalique devient plus difficile.
La mise au point d'un vaccin sur la base d'une immunoglobuline G fait l'objet de recherches. Ce vaccin devrait stimuler des anticorps se liant spécifiquement à la nicotine, et par suite empêchant son passage à travers la barrière hémato-encéphalique.
Ondes électromagnétiques (téléphones mobiles)
Les effets adverses sur la santé des rayonnements électromagnétiques dans le domaine du MHz au GHz à haute densité d'énergie sont bien connus. C'est avec eux que l'on fait cuire la nourriture au four à micro-ondes. Cependant, les effets des rayonnements de densité d'énergie bien inférieure, comme en téléphonie ou application multimédia mobile, sont sujets à controverses. Les effets spécifiques sur la barrière hémato-encéphalique sont un domaine d'incertitude.
À haute densité d'énergie du rayonnement électromagnétique, on observe un échauffement significatif du tissu corporel. Dans le crâne, ce réchauffement pourrait influencer la barrière hémato-encéphalique et la rendre plus perméable. On observe ce genre d'effets de réchauffement sur des organes périphériques. Dans les circonstances de la téléphonie mobile, le cerveau s'échauffe au maximum de 0,1 K (15 minutes de conversation à puissance d'émission maximum). Un bain chaud ou un travail corporel fatigant peuvent échauffer plus fort le cerveau sans danger. Des études scientifiques datant du début des années 1990, en particulier dans le groupe du neurochirurgien suédois Leif G. Salford de l'université de Lund,rapportent une ouverture de la barrière hémato-encéphalique dans le domaine non thermique avec des fréquences GSM.
D'autres groupes de travail ne confirment pas les résultats de Salford, certains remettant en cause la méthode utilisée.
Diagnostics en médecine humaine
IRM renforcée par produit de contraste

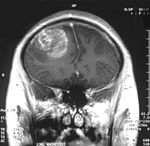

Le premier produit de contraste mis au point pour l'IRM est le gadolinium (Gd). En raison de sa toxicité, il faut l’emballer (le chélater) dans une molécule de DTPA. On a ainsi obtenu en 1984 le Gd-DTPA, qui avait le potentiel pour obtenir des IRM renforcés pour le diagnostic de lésions locales de la barrière hémato-encéphalique. La molécule de Gd-DTPA est très polaire, et par conséquent bien trop hydrophile pour traverser une barrière hémato-encéphalique saine. Les modifications des jonctions serrées, comme celles qui peuvent par exemple être provoquées par un glioblastome, permettent le transport paracellulaire de ce produit de contraste dans le tissu cérébral. Là, il renforce le contraste, par interaction avec les protons de l'eau environnante, et rend visibles les défauts de la barrière hémato-encéphalique. Comme ce sont les vaisseaux responsables de l'alimentation de la tumeur qui sont touchés, dans son voisinage immédiat, on peut en apprécier l'extension.
Au cas d'un accident vasculaire cérébral aigu, le dommage à la barrière hémato-encéphalique peut être soumis au diagnostic de la même manière par IRM renforcée par produit de contraste.
Par la détermination du temps de relaxation, la quantité de Gd-DTPA dans le tissu cérébral peut être quantifiée.
Autres procédés d'imagerie
Au moyen de traceurs marqués par un élément radioactif, qui ne passent pas normalement à travers la barrière hémato-encéphalique, on peut aussi entreprendre des recherches sur le fonctionnement de celle-ci chez l'homme. Pour cela, on peut en principe utiliser la tomographie d'émission monophotonique (TEMP, ou en anglais SPECT), ou la tomographie par émission de positons (TEP, ou en anglais PET).
Par exemple, chez des patients victimes d'un accident vasculaire cérébral aigu, on peut montrer une augmentation de l'absorption du 99mTc chélaté par l'hexa-methyl-propylene-amine-oxime (HMPAO).
Au moyen de la tomodensitométrie, on peut aussi quantifier les défauts de la barrière hémato-encéphalique par la diffusion de produits de contraste appropriés hors des capillaires.
Histoire de la découverte de la barrière hémato-encéphalique

La première preuve d'existence pour la barrière hémato-encéphalique vient du chimiste allemand Paul Ehrlich. En 1885, il constata qu'après injection de colorants vitaux solubles dans l'eau dans la circulation sanguine de rats, tous les organes étaient colorés, sauf le cerveau et la moelle épinière.
En 1904, il en tira une conclusion fausse, c'est-à-dire que la cause de cette découverte était une faible affinité du tissu cérébral pour le colorant injecté.
En 1909, Edwin Goldmann, un ancien collaborateur de Paul Ehrlich, injecte par intraveineuse le colorant synthétisé cinq ans auparavant par Ehrlich, le bleu de trypan, un colorant azoïque. Là-dessus, il remarque que le plexus choroideus, contrairement au tissu cérébral qui l'entoure, est coloré de façon marquée. En 1913, il injecte la même substance directement dans le liquide cérébrospinal de chiens et lapins. Goldmann en conclut que le liquide cérébrospinal et le plexus choroideus ont une fonction importante dans le transport des nutriments du système nerveux central. De plus, il soupçonne une fonction de barrière contre les substances neurotoxiques.
En 1898, Arthur Biedl et Rudolf Kraus mènent des expériences avec l’acide gallique. Ce composé se révèle non toxique par application dans la circulation générale. Mais son injection dans le cerveau s'avère neurotoxique, avec des réactions pouvant aller jusqu'au coma.
Max Lewandowsky utilise en 1900 pour des expériences semblables le ferrocyanure de potassium et arrive à des conclusions semblables à celles de Biedl et Kraus. Lewandowsky utilise à cette occasion pour la première fois le concept de « barrière hémato-encéphalique ».

En 1890, Charles Smart Roy et le futur prix Nobel de médecine Charles Scott Sherrington postulent que le cerveau possède un mécanisme intrinsèque pour faire correspondre l'irrigation vasculaire aux variations locales de l'activité :
« Le cerveau possède un mécanisme intrinsèque par lequel l'apport vasculaire peut être varié localement en correspondance avec les variations locales d'activité fonctionnelle. »

Lina Stern née le 26 août 1878 et morte 7 mars 1968 à Moscou, femme médecin et biochimiste soviétique, première membre féminine de l'académie des sciences de Russie, a apporté de réelles contributions à la recherche sur la barrière hémato-encéphalique, qu'elle désigna comme telle en 1921.
La différence entre la barrière hémato-encéphalique et la barrière sang - liquide cérébrospinal fut prise en compte dans les années 1930 par Friedrich Karl Walter et Hugo Spatz. Ils ont posé que le flux de liquide cérébrospinal était par lui-même insuffisant pour assurer l'échange de gaz du système nerveux central.
Bien que les expériences de Goldmann et Ehrlich eussent indiqué l'existence d'une barrière entre la circulation sanguine et le système nerveux central, ce n'est que dans les années 1960 que les derniers doutes concernant son existence ont été dissipés. Un point critique dans l'expérience de Goldmann consistait en ce que le sang et le liquide cérébro-spinal, les deux liquides dans lesquels il avait injecté des colorants, diffèrent considérablement, ce qui pouvait influer sur le comportement de la diffusion et l'affinité pour le tissu nerveux. La compréhension a été rendue encore plus difficile par la trouvaille expérimentale que les colorants azoïques basiques coloraient le tissu nerveux, donc franchissaient la barrière, tandis que les colorants acides ne le faisaient pas. Ulrich Friedemann en conclut que c'étaient les propriétés électrochimiques des colorants qui en étaient responsables : les capillaires cérébraux étaient perméables aux substances neutres ou de pH supérieur au sang, et imperméables aux autres. Mais par la suite, quand un grand nombre de substances furent testées pour leur capacité à franchir la barrière hémato-encéphalique, cette hypothèse se révéla insuffisante. Dans les modèles d'explication suivants, on introduisit et soumit à discussion tout une série de paramètres, comme la masse molaire, la taille de la molécule, les affinités de liaison, les constantes de dissociation, le caractère lipophile, la charge électrique, et leurs diverses combinaisons.
La compréhension actuelle de la structure de base de la barrière hémato-encéphalique est fondée sur des vues au microscope électronique de cerveaux de souris, que l'on arriva à faire à la fin des années 1960. Thomas S. Reese et Morris J. Karnovsky ont injecté à leurs sujets animaux pendant leurs expériences de la peroxydase de raifort (HRP) par voie intraveineuse. Ils n'ont trouvé l'enzyme, au microscope électronique, que dans la lumière des capillaires et dans des vésicules micropinocytaires au sein des cellules endothéliales. À l'extérieur des endothéliums, dans la matrice extracellulaire, ils n'ont pas trouvé de peroxydase. Ils en ont conclu que les jonctions serrées entre les cellules endothéliales empêchent le passage vers le cerveau.
Voir aussi
Bibliographie
- (de) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en allemand intitulé « Blut-Hirn-Schranke » (voir la liste des auteurs).
- Extraits du cours de PCEM1 du Pr Bertrand Bloch (PU-PH) sur le tissu nerveux, Université Victor Segalen, Bordeaux 2.
- (en) D. Kobiler, Blood-brain Barrier., Springer Verlag, (ISBN 0-306-46708-9)
- (en) A. G. De Boer et W. Sutanto, Drug Transport Across the Blood-brain Barrier., CRC Press, , 216 p. (ISBN 90-5702-032-7, présentation en ligne)
- (en) W. M. Pardridge, Introduction to the Blood-brain Barrier., Cambridge University Press, (ISBN 0-521-58124-9)
- (en) E. M. Taylor, Efflux Transporters and the Blood-brain Barrier., Nova Publishers, , 247 p. (ISBN 1-59454-625-8, présentation en ligne)
- (en) D. J. Begley, The Blood-brain Barrier and Drug Delivery to the CNS., Informa Health Care, (ISBN 0-8247-0394-4)
- (en) E. de Vries et A. Prat, The Blood-brain Barrier and Its Microenvironment., Taylor & Francis, (ISBN 0-8493-9892-4)
- (en) M. Bradbury, The Concept of a Blood-Brain Barrier., Wiley-Interscience, (ISBN 0-471-99688-2)
- (de) P. Ramge, Untersuchungen zur Überwindung der Blut-Hirn-Schranke mit Hilfe von Nanopartikeln., Shaker Verlag, (ISBN 3-8265-4974-0)
- (de) Peter Uwe Brenner, « Die Struktur der Blut-Hirn- und der Blut-Liquor-Schranke. – Eine Literaturstudie – », Thèse de doctorat, Ludwig Maximilans Universität München, (lire en ligne)