
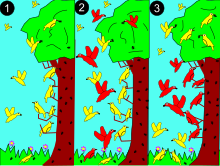
2: une espèce plus grande (rouge) entre en compétition pour les ressources alimentaires.
3: la rouge plus efficace dans l'utilisation des ressources des strates moyennes, exclut la jaune qui est refoulée dans deux nouvelles niches, la strate supérieure et inférieure des arbres
Ce principe est bien mis en évidence chez cinq espèces de fauvettes consommant des insectes dans une partie différente des épicéas : elles évitent la compétition par une ségrégation spatiale de leurs niches.
En écologie fondamentale, et plus particulièrement en biologie des populations, le principe de Gause (appelé aussi principe d'exclusion réciproque, principe d'exclusion compétitive, principe d'exclusion de compétition ou principe de Volterra-Gause) est un principe de la dynamique des populations stipulant que deux populations ne peuvent subsister sur la même niche écologique. L'exclusion compétitive conduit à la différenciation de niche.
Historique
Le principe est habituellement attribué à Georgy Gause, bien que celui-ci ne l'ait jamais énoncé, et que le principe soit déjà présent chez Darwin.
L'énoncé du principe
Il stipule que les populations de deux espèces ayant des exigences écologiques identiques, c'est-à-dire exploitant une ressource limitante unique, ne peuvent coexister indéfiniment dans un milieu stable et homogène. L'écologue Garrett Hardin précise cette hypothèse en 1960 sous la forme du « principe d'exclusion par compétition » qui prévoit que l'espèce qui est la plus efficace dans l'utilisation d'une ressource limitée du milieu finit par éliminer la ou les espèces voisines, moins aptes à cette utilisation. Le refoulement de ces dernières s'effectue moins par attaque directe que par privation des facteurs vitaux objets de la compétition (préférences stationnelles, besoins en eau et en nutriments, etc., chez les plantes ; choix de la nourriture, lieux d'alimentation, choix des abris, des lieux de reproduction, etc., chez les animaux). Le refoulement impose au vaincu de nouvelles conditions de vie en occupant une nouvelle niche.
Ce principe revient à dire que deux espèces ne peuvent partager la même niche écologique (ensemble des exigences écologiques). C'est d'ailleurs grâce à ce principe que cette notion fut introduite.
Le corollaire du principe de Gause consiste à dire que si deux espèces écologiquement semblables coexistent, c'est qu'elles ont nécessairement réalisé une différenciation de niche.
L'expérience originelle de Gause (1934)
L'expérience originelle de Gause, réalisée sur des paramécies, consistait à quantifier une population de Paramecium caudatum et une population de Paramecium aurelia cultivées dans le même milieu nutritif. L'expérience montre que Paramecium aurelia est l'espèce la plus compétitive et élimine l'autre en quelques jours.
Portée et limites
De nombreux autres résultats expérimentaux et observations de terrain ont depuis montré que ce principe s'étendait à de nombreux domaines, s'appliquant autant aux animaux qu'aux végétaux. Pour beaucoup d'auteurs, il existe cependant des cas limites voire des exceptions à ce principe, deux espèces sympatriques parfaitement compétitives pouvant dans certains cas coexister.
L'une des limites du principe est qu'il oscille entre principe a priori (des espèces coexistant doivent avoir des niches différentes, même si on ne voit pas les différences) et principe a posteriori (on vérifie bien que les espèces coexistant ont des niches différentes).
L'hyperdiversité dans les forêts tropicales humides ne peut s'expliquer par ce principe. L'effet Janzen–Connell propose que certaines niches sont délimitées par des rétroactions négatives dans lesquelles des plants adultes, via des interactions intraspécifiques, inhibent le développement de jeunes plants de la même espèce à proximité d'eux. Quand une espèce s'installe, elle attire des phytophages (granivores, frugivores), des parasites (vecteurs d'infection, comme les spores) et des pathogènes spécifiques, limitant l'installation des individus suivants de la même espèce et favorisant celle d'autres espèces dans la même niche écologique. L'hypothèse de Daniel Janzen (en) et Joseph Connell (en), émise en 1970, pourrait aussi s'exercer dans les prairies par l'intermédiaire des microorganismes pathogènes du sol et sur le processus de succession écologique (les pathogènes entraînent un déclin de vigueur d'une espèce, remplacée par une autre espèce compétitive qui n'a pas les mêmes pathogènes).
Voir aussi
Bibliographie
- Duquet Marc, 1993. Glossaire d'écologie fondamentale, éd. Fernand Nathan, 127 p. (ISBN 2-09-190538-0)
- Depotte C., Djegham Y., Noël G., Verhaeghe J.-C., 2003. Mathématique & biologie, une expérience pluridisciplinaire. éd. de Boeck, 208 pages, p. 68. (ISBN 2-8041-4314-7)
- De Robert, Ricklefs, Gary, Miller, 2005. Écologie. éd. de Boeck, 858 pages, p. 407. (ISBN 2-7445-0145-X)
- Dajoz R., 2006. Précis d'écologie. Dunod éd., 640 p. (ISBN 978-2-10-049627-3)
- Gause G. F., 1935. Vérifications expérimentales de la théorie mathématique de la lutte pour la vie. Hermann et Cie, éditeurs, Paris, France.
- glossaire d'écologie de R. Dajoz (complément à Précis d'écologie, 2006) [1]
Articles connexes
- Compétition (biologie)
- Niche écologique
- Sélection naturelle
- Equations de compétition de Lotka-Volterra